

Caliciviridae Other Than Noroviruses
- PMID: 30901945
- PMCID: PMC6466229
- DOI: 10.3390/v11030286
Caliciviridae Other Than Noroviruses
Abstract
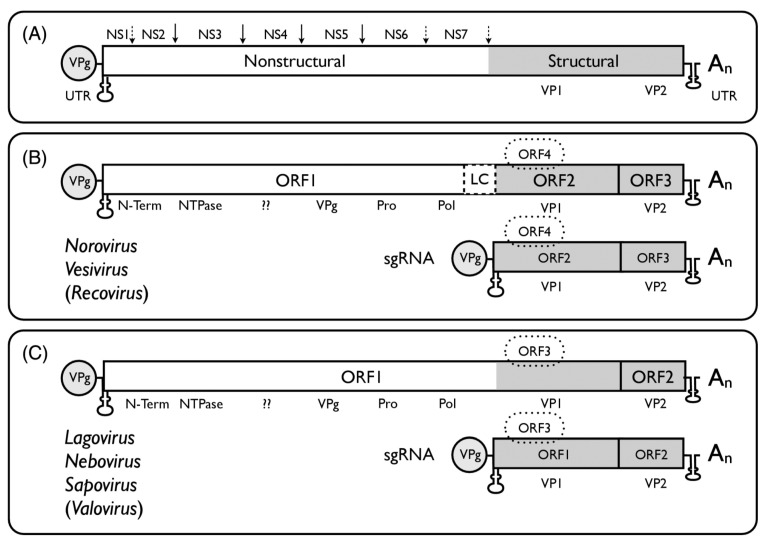
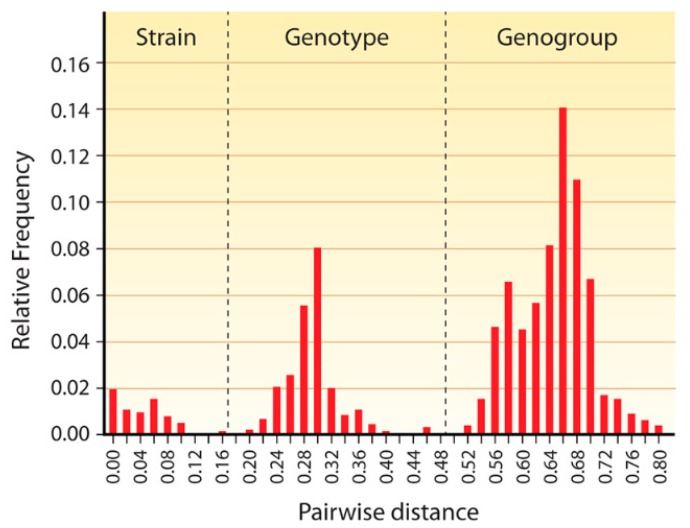
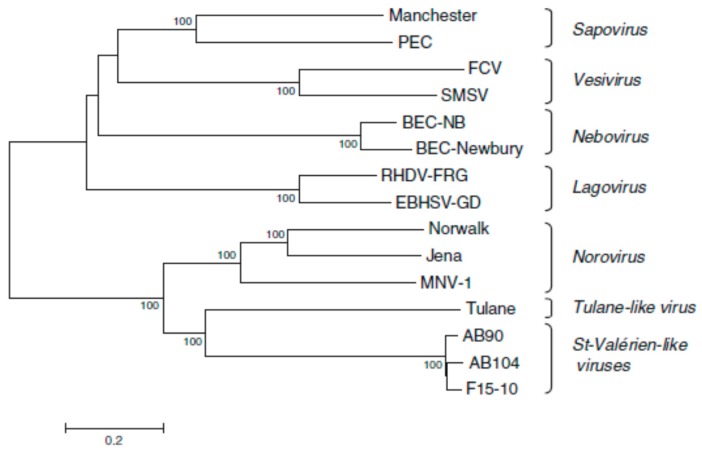
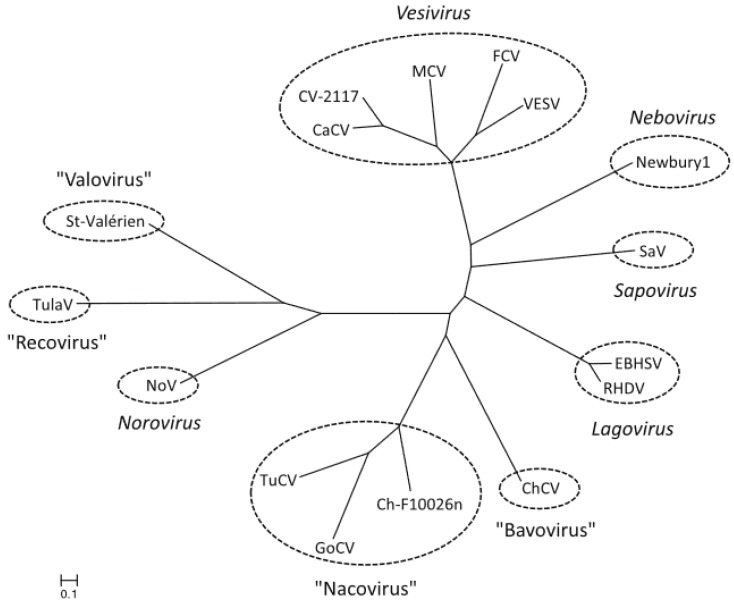
Besides noroviruses, the Caliciviridae family comprises four other accepted genera: Sapovirus, Lagovirus, Vesivirus, and Nebovirus. There are six new genera proposed: Recovirus, Valovirus, Bavovirus, Nacovirus, Minovirus, and Salovirus. All Caliciviridae have closely related genome structures, but are genetically and antigenically highly diverse and infect a wide range of mammalian host species including humans. Recombination in nature is not infrequent for most of the Caliciviridae, contributing to their diversity. Sapovirus infections cause diarrhoea in pigs, humans and other mammalian hosts. Lagovirus infections cause systemic haemorrhagic disease in rabbits and hares, and vesivirus infections lead to lung disease in cats, vesicular disease in swine, and exanthema and diseases of the reproductive system in large sea mammals. Neboviruses are an enteric pathogen of cattle, differing from bovine norovirus. At present, only a few selected caliciviruses can be propagated in cell culture (permanent cell lines or enteroids), and for most of the cultivatable caliciviruses helper virus-free, plasmid only-based reverse genetics systems have been established. The replication cycles of the caliciviruses are similar as far as they have been explored: viruses interact with a multitude of cell surface attachment factors (glycans) and co-receptors (proteins) for adsorption and penetration, use cellular membranes for the formation of replication complexes and have developed mechanisms to circumvent innate immune responses. Vaccines have been developed against lagoviruses and vesiviruses, and are under development against human noroviruses.
Keywords: Bavovirus; Lagovirus; Nebovirus; Norovirus; Recovirus; Sapovirus; Valovirus; Vesivirus; animal models; enteroids; reverse genetics.
Conflict of interest statement
The author declares no conflict of interest.
Figures
Similar articles
-
ICTV Virus Taxonomy Profile: Caliciviridae.J Gen Virol. 2019 Nov;100(11):1469-1470. doi: 10.1099/jgv.0.001332. J Gen Virol. 2019. PMID: 31573467 Free PMC article.
-
Calicivirus Non-structural Proteins: Potential Functions in Replication and Host Cell Manipulation.Front Microbiol. 2021 Jul 14;12:712710. doi: 10.3389/fmicb.2021.712710. eCollection 2021. Front Microbiol. 2021. PMID: 34335548 Free PMC article. Review.
-
Bat Caliciviruses and Human Noroviruses Are Antigenically Similar and Have Overlapping Histo-Blood Group Antigen Binding Profiles.mBio. 2018 May 22;9(3):e00869-18. doi: 10.1128/mBio.00869-18. mBio. 2018. PMID: 29789360 Free PMC article.
-
Porcine enteric caliciviruses: genetic and antigenic relatedness to human caliciviruses, diagnosis and epidemiology.Vaccine. 2007 Jul 26;25(30):5453-66. doi: 10.1016/j.vaccine.2006.12.032. Epub 2006 Dec 29. Vaccine. 2007. PMID: 17234307 Free PMC article. Review.
-
Animal noroviruses.Vet J. 2008 Oct;178(1):32-45. doi: 10.1016/j.tvjl.2007.11.012. Epub 2008 Feb 21. Vet J. 2008. PMID: 18294883 Review.
Cited by
-
Genetic Diversity of Sapoviruses among Inpatients in Germany, 2008-2018.Viruses. 2019 Aug 7;11(8):726. doi: 10.3390/v11080726. Viruses. 2019. PMID: 31394867 Free PMC article.
-
Noroviruses-The State of the Art, Nearly Fifty Years after Their Initial Discovery.Viruses. 2021 Aug 4;13(8):1541. doi: 10.3390/v13081541. Viruses. 2021. PMID: 34452406 Free PMC article. Review.
-
Isolation of a rhesus calicivirus that can replicate in human cells.Virology. 2023 May;582:83-89. doi: 10.1016/j.virol.2023.03.012. Epub 2023 Apr 2. Virology. 2023. PMID: 37031656 Free PMC article.
-
Clinical Profiles of Childhood Astrovirus-, Sapovirus-, and Norovirus-Associated Acute Gastroenteritis in Pediatric Emergency Departments in Alberta, 2014-2018.J Infect Dis. 2022 Feb 15;225(4):723-732. doi: 10.1093/infdis/jiab429. J Infect Dis. 2022. PMID: 34432027 Free PMC article.
-
Bile acids and ceramide overcome the entry restriction for GII.3 human norovirus replication in human intestinal enteroids.Proc Natl Acad Sci U S A. 2020 Jan 21;117(3):1700-1710. doi: 10.1073/pnas.1910138117. Epub 2020 Jan 2. Proc Natl Acad Sci U S A. 2020. PMID: 31896578 Free PMC article.
References
-
- Wobus C.E., Karst S.M., Thackray L.B., Chang K.O., Sosnovtsev S.V., Belliot G., Krug A., Mackenzie J.M., Green K.Y., Virgin H.W. Replication of Norovirus in cell culture reveals a tropism for dendritic cells and macrophages. PLoS Biol. 2004;2:e432. doi: 10.1371/journal.pbio.0020432. - DOI - PMC - PubMed
-
- Ward V.K., McCormick C.J., Clarke I.N., Salim O., Wobus C.E., Thackray L.B., Virgin H.W., 4th, Lambden P.R. Recovery of infectious murine norovirus using pol II-driven expression of full-length cDNA. Proc. Natl. Acad. Sci. USA. 2007;104:11050–11055. doi: 10.1073/pnas.0700336104. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
