

Small-Molecule Agonists of Ae. aegypti Neuropeptide Y Receptor Block Mosquito Biting
- PMID: 30735632
- PMCID: PMC6369589
- DOI: 10.1016/j.cell.2018.12.004
Small-Molecule Agonists of Ae. aegypti Neuropeptide Y Receptor Block Mosquito Biting
Abstract
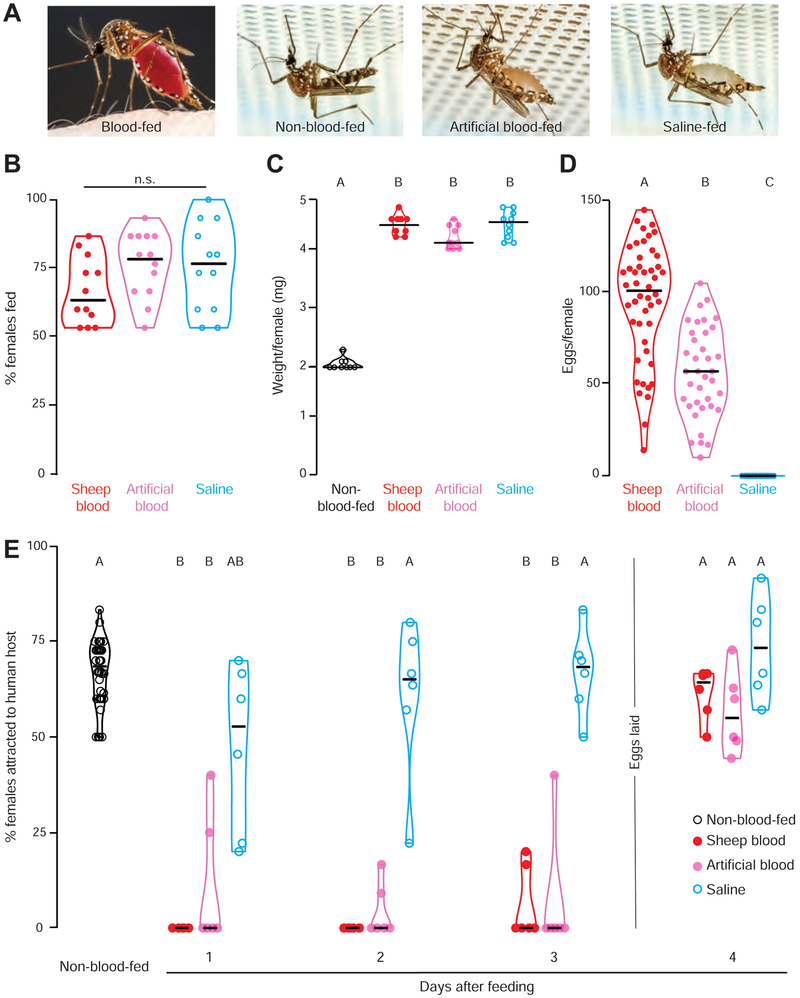
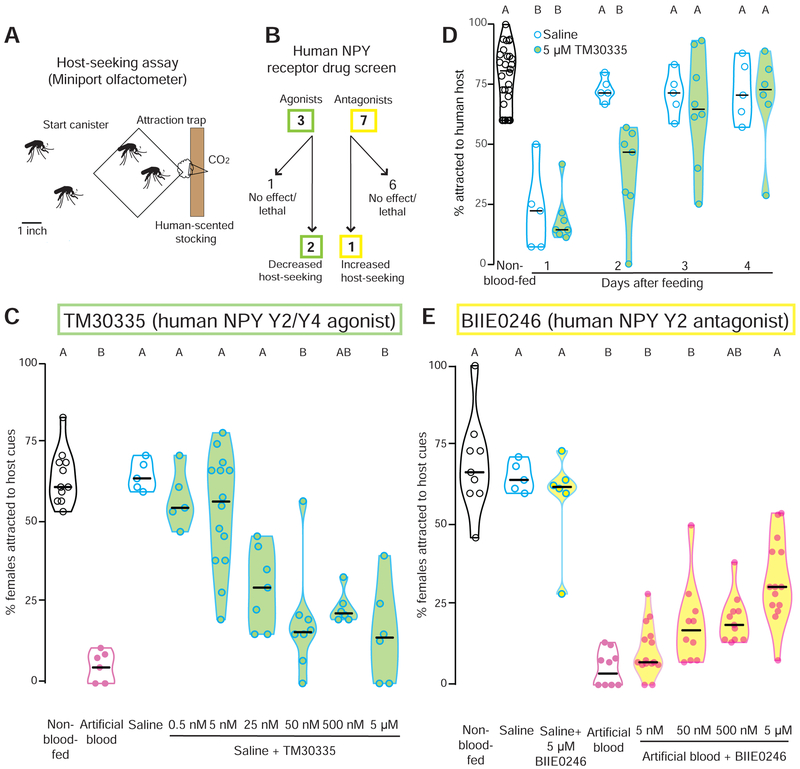
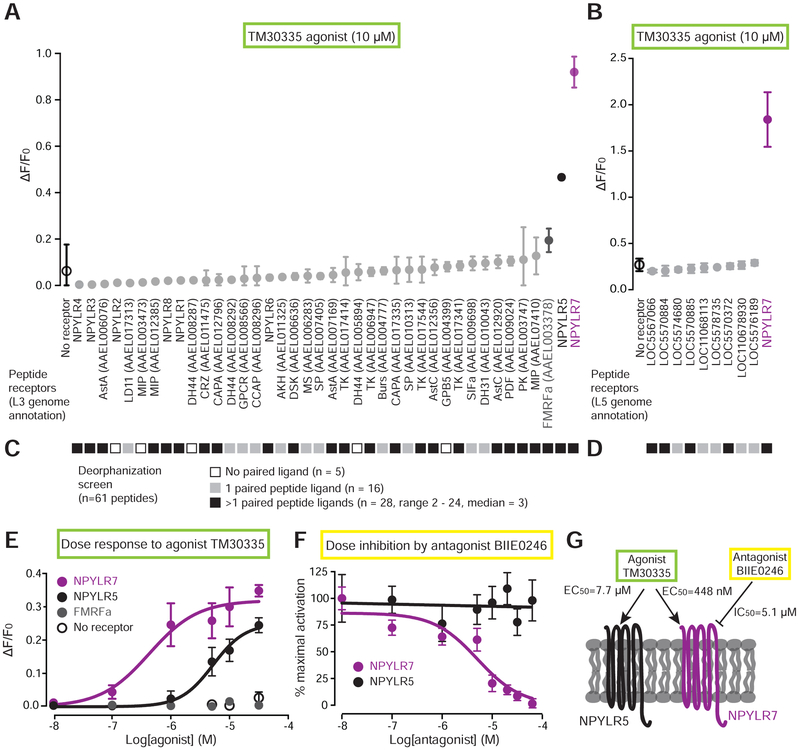
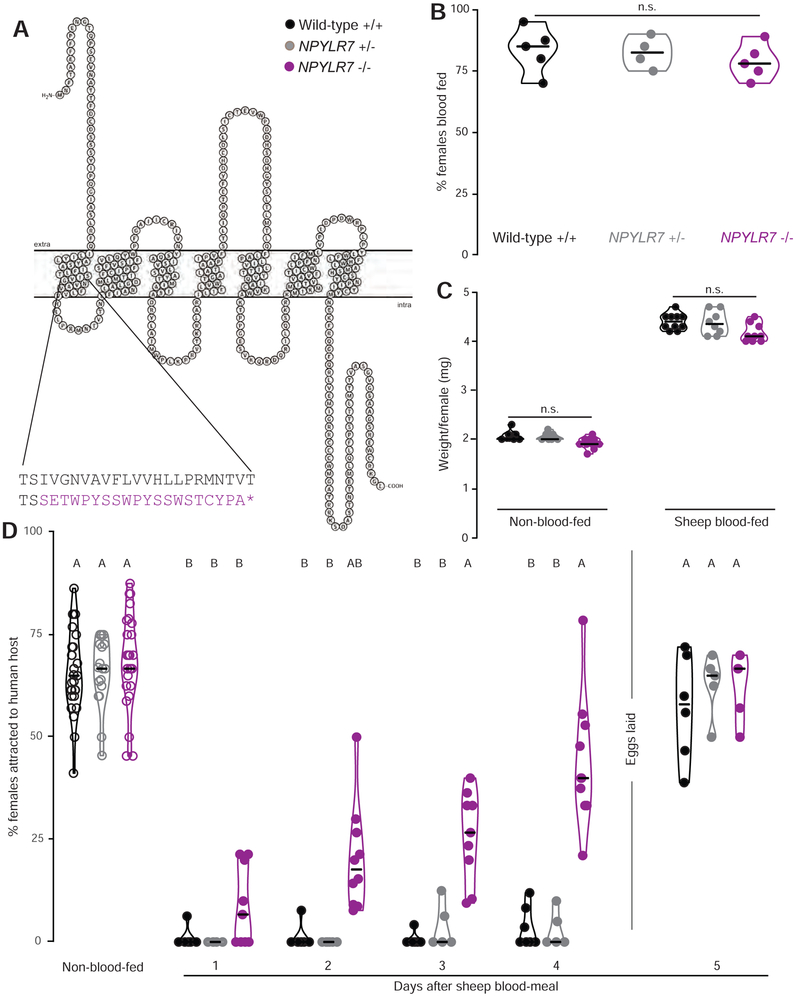
Female Aedes aegypti mosquitoes bite humans to obtain blood to develop their eggs. Remarkably, their strong attraction to humans is suppressed for days after the blood meal by an unknown mechanism. We investigated a role for neuropeptide Y (NPY)-related signaling in long-term behavioral suppression and discovered that drugs targeting human NPY receptors modulate mosquito host-seeking. In a screen of all 49 predicted Ae. aegypti peptide receptors, we identified NPY-like receptor 7 (NPYLR7) as the sole target of these drugs. To obtain small-molecule agonists selective for NPYLR7, we performed a high-throughput cell-based assay of 265,211 compounds and isolated six highly selective NPYLR7 agonists that inhibit mosquito attraction to humans. NPYLR7 CRISPR-Cas9 null mutants are defective in behavioral suppression and resistant to these drugs. Finally, we show that these drugs can inhibit biting and blood-feeding on a live host, suggesting a novel approach to control infectious disease transmission by controlling mosquito behavior. VIDEO ABSTRACT.
Keywords: Aedes aegypti; CRISPR-Cas9; Zika; blood-feeding; dengue; feeding; high-throughput small-molecule screen; host-seeking behavior; mosquito; neuropeptide Y.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures

Comment in
-
The Perfect Appetizer: A Pharmacological Strategy for a Non-biting Mosquito.Cell. 2019 Feb 7;176(4):679-681. doi: 10.1016/j.cell.2019.01.032. Cell. 2019. PMID: 30735629
-
Mosquitoes on a Diet Reduce Those Pesky Bites.Trends Parasitol. 2019 May;35(5):335-336. doi: 10.1016/j.pt.2019.03.003. Epub 2019 Mar 23. Trends Parasitol. 2019. PMID: 30910491
-
Diet Drugs Trick Mosquitoes into Feeling Full.Trends Pharmacol Sci. 2019 Jul;40(7):449-451. doi: 10.1016/j.tips.2019.04.016. Epub 2019 May 20. Trends Pharmacol Sci. 2019. PMID: 31122765
Similar articles
-
Next-generation neuropeptide Y receptor small-molecule agonists inhibit mosquito-biting behavior.Parasit Vectors. 2024 Jun 28;17(1):276. doi: 10.1186/s13071-024-06347-w. Parasit Vectors. 2024. PMID: 38937807 Free PMC article.
-
Diet Drugs Trick Mosquitoes into Feeling Full.Trends Pharmacol Sci. 2019 Jul;40(7):449-451. doi: 10.1016/j.tips.2019.04.016. Epub 2019 May 20. Trends Pharmacol Sci. 2019. PMID: 31122765
-
The Perfect Appetizer: A Pharmacological Strategy for a Non-biting Mosquito.Cell. 2019 Feb 7;176(4):679-681. doi: 10.1016/j.cell.2019.01.032. Cell. 2019. PMID: 30735629
-
Similar Neural Pathways Control Foraging in Mosquitoes and Worms.mBio. 2019 Apr 30;10(2):e00656-19. doi: 10.1128/mBio.00656-19. mBio. 2019. PMID: 31040241 Free PMC article. Review.
-
Mosquito Host-Seeking Regulation: Targets for Behavioral Control.Trends Parasitol. 2019 Sep;35(9):704-714. doi: 10.1016/j.pt.2019.06.010. Epub 2019 Jul 17. Trends Parasitol. 2019. PMID: 31326312 Review.
Cited by
-
Updated Role of Neuropeptide Y in Nicotine-Induced Endothelial Dysfunction and Atherosclerosis.Front Cardiovasc Med. 2021 Feb 23;8:630968. doi: 10.3389/fcvm.2021.630968. eCollection 2021. Front Cardiovasc Med. 2021. PMID: 33708805 Free PMC article. Review.
-
QnAs with Leslie B. Vosshall.Proc Natl Acad Sci U S A. 2020 Jul 14;117(28):16100-16103. doi: 10.1073/pnas.2011073117. Epub 2020 Jun 29. Proc Natl Acad Sci U S A. 2020. PMID: 32601194 Free PMC article. No abstract available.
-
Targeting Aedes aegypti Metabolism with Next-Generation Insecticides.Viruses. 2023 Feb 8;15(2):469. doi: 10.3390/v15020469. Viruses. 2023. PMID: 36851683 Free PMC article. Review.
-
Nano-pesticides: the lunch-box principle-deadly goodies (semio-chemical functionalised nanoparticles that deliver pesticide only to target species).J Nanobiotechnology. 2022 Jan 4;20(1):13. doi: 10.1186/s12951-021-01216-5. J Nanobiotechnology. 2022. PMID: 34983544 Free PMC article.
-
Bidirectional Microbiome-Gut-Brain-Axis Communication Influences Metabolic Switch-Associated Responses in the Mosquito Anopheles culicifacies.Cells. 2022 May 31;11(11):1798. doi: 10.3390/cells11111798. Cells. 2022. PMID: 35681493 Free PMC article.
References
-
- Anderson JF, and Magnarelli LA (2008). Biology of ticks. Infectious disease clinics of North America 22, 195–215. - PubMed
-
- Batterham RL, Cowley MA, Small CJ, Herzog H, Cohen MA, Dakin CL, Wren AM, Brynes AE, Low MJ, Ghatei MA, et al. (2002). Gut hormone PYY(3-36) physiologically inhibits food intake. Nature 418, 650–654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
