

Allosteric Inhibition of Ubiquitin-like Modifications by a Class of Inhibitor of SUMO-Activating Enzyme
- PMID: 30581133
- PMCID: PMC6524651
- DOI: 10.1016/j.chembiol.2018.10.026
Allosteric Inhibition of Ubiquitin-like Modifications by a Class of Inhibitor of SUMO-Activating Enzyme
Abstract
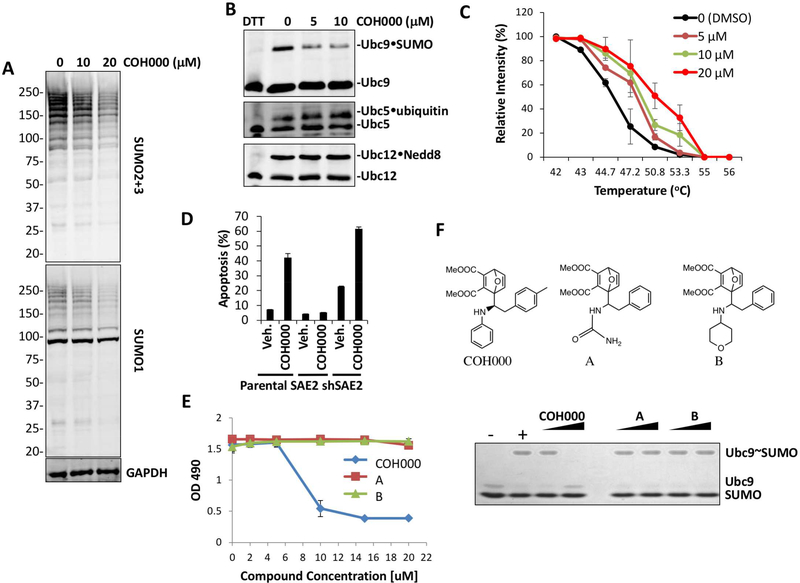
Ubiquitin-like (Ubl) post-translational modifications are potential targets for therapeutics. However, the only known mechanism for inhibiting a Ubl-activating enzyme is through targeting its ATP-binding site. Here we identify an allosteric inhibitory site in the small ubiquitin-like modifier (SUMO)-activating enzyme (E1). This site was unexpected because both it and analogous sites are deeply buried in all previously solved structures of E1s of ubiquitin-like modifiers (Ubl). The inhibitor not only suppresses SUMO E1 activity, but also enhances its degradation in vivo, presumably due to a conformational change induced by the compound. In addition, the lead compound increased the expression of miR-34b and reduced c-Myc levels in lymphoma and colorectal cancer cell lines and a colorectal cancer xenograft mouse model. Identification of this first-in-class inhibitor of SUMO E1 is a major advance in modulating Ubl modifications for therapeutic aims.
Keywords: E1; KRas; SUMO; activating enzyme; allosteric inhibitor; c-Myc; cancer; covalent inhibitor; therapeutics; ubiquitin-like modification.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interests
S.X.O. is an employee and shareholder of SUMO Biosciences, Inc. Y.C. is a founder of SUMO Biosciences, Inc. and a member of its advisory board. Other authors declare no competing interests.
Figures




Comment in
-
Discovery of a First-In-Class Covalent Allosteric Inhibitor of SUMO E1 Activating Enzyme.Cell Chem Biol. 2019 Feb 21;26(2):153-155. doi: 10.1016/j.chembiol.2019.02.006. Cell Chem Biol. 2019. PMID: 30794786
Similar articles
-
Molecular mechanism of a covalent allosteric inhibitor of SUMO E1 activating enzyme.Nat Commun. 2018 Dec 4;9(1):5145. doi: 10.1038/s41467-018-07015-1. Nat Commun. 2018. PMID: 30514846 Free PMC article.
-
Designed semisynthetic protein inhibitors of Ub/Ubl E1 activating enzymes.J Am Chem Soc. 2010 Feb 17;132(6):1748-9. doi: 10.1021/ja9088549. J Am Chem Soc. 2010. PMID: 20099854 Free PMC article.
-
Discovery of TAK-981, a First-in-Class Inhibitor of SUMO-Activating Enzyme for the Treatment of Cancer.J Med Chem. 2021 Mar 11;64(5):2501-2520. doi: 10.1021/acs.jmedchem.0c01491. Epub 2021 Feb 25. J Med Chem. 2021. PMID: 33631934
-
Regulating the Master Regulator: Controlling Ubiquitination by Thinking Outside the Active Site.J Med Chem. 2018 Jan 25;61(2):405-421. doi: 10.1021/acs.jmedchem.6b01346. Epub 2017 Feb 28. J Med Chem. 2018. PMID: 28076680 Review.
-
The role of allostery in the ubiquitin-proteasome system.Crit Rev Biochem Mol Biol. 2013 Mar-Apr;48(2):89-97. doi: 10.3109/10409238.2012.742856. Epub 2012 Dec 13. Crit Rev Biochem Mol Biol. 2013. PMID: 23234564 Free PMC article. Review.
Cited by
-
Targeting SUMOylation dependency in human cancer stem cells through a unique SAE2 motif revealed by chemical genomics.Cell Chem Biol. 2021 Oct 21;28(10):1394-1406.e10. doi: 10.1016/j.chembiol.2021.04.014. Epub 2021 May 11. Cell Chem Biol. 2021. PMID: 33979648 Free PMC article.
-
SUMO: From Bench to Bedside.Physiol Rev. 2020 Oct 1;100(4):1599-1619. doi: 10.1152/physrev.00025.2019. Physiol Rev. 2020. PMID: 32666886 Free PMC article. Review.
-
Critical Non-Covalent Binding Intermediate for an Allosteric Covalent Inhibitor of SUMO E1.J Phys Chem Lett. 2023 Mar 23;14(11):2792-2799. doi: 10.1021/acs.jpclett.3c00253. Epub 2023 Mar 10. J Phys Chem Lett. 2023. PMID: 36898086 Free PMC article.
-
Poly-SUMO-2/3 chain modification of Nuf2 facilitates CENP-E kinetochore localization and chromosome congression during mitosis.Cell Cycle. 2021 May;20(9):855-873. doi: 10.1080/15384101.2021.1907509. Epub 2021 Apr 28. Cell Cycle. 2021. PMID: 33910471 Free PMC article.
-
SENP3 affects the expression of PYCR1 to promote bladder cancer proliferation and EMT transformation by deSUMOylation of STAT3.Aging (Albany NY). 2022 Oct 11;14(19):8032-8045. doi: 10.18632/aging.204333. Epub 2022 Oct 11. Aging (Albany NY). 2022. PMID: 36227136 Free PMC article.
References
-
- Boggio R, Colombo R, Hay RT, Draetta GF, and Chiocca S (2004). A mechanism for inhibiting the SUMO pathway. Molecular cell 16, 549–561. - PubMed
-
- Bohnsack RN, and Haas AL (2003). Conservation in the mechanism of Nedd8 activation by the human AppBp1-Uba3 heterodimer. J Biol Chem 278, 26823–26830. - PubMed
-
- Brownell JE, Sintchak MD, Gavin JM, Liao H, Bruzzese FJ, Bump NJ, Soucy TA, Milhollen MA, Yang X, Burkhardt AL, et al. (2010). Substrate-assisted inhibition of ubiquitin-like protein-activating enzymes: the NEDD8 E1 inhibitor MLN4924 forms a NEDD8-AMP mimetic in situ. Molecular cell 37, 102–111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
