

Functional and Physical Interaction between the Arf Activator GBF1 and Hepatitis C Virus NS3 Protein
- PMID: 30567983
- PMCID: PMC6401423
- DOI: 10.1128/JVI.01459-18
Functional and Physical Interaction between the Arf Activator GBF1 and Hepatitis C Virus NS3 Protein
Abstract
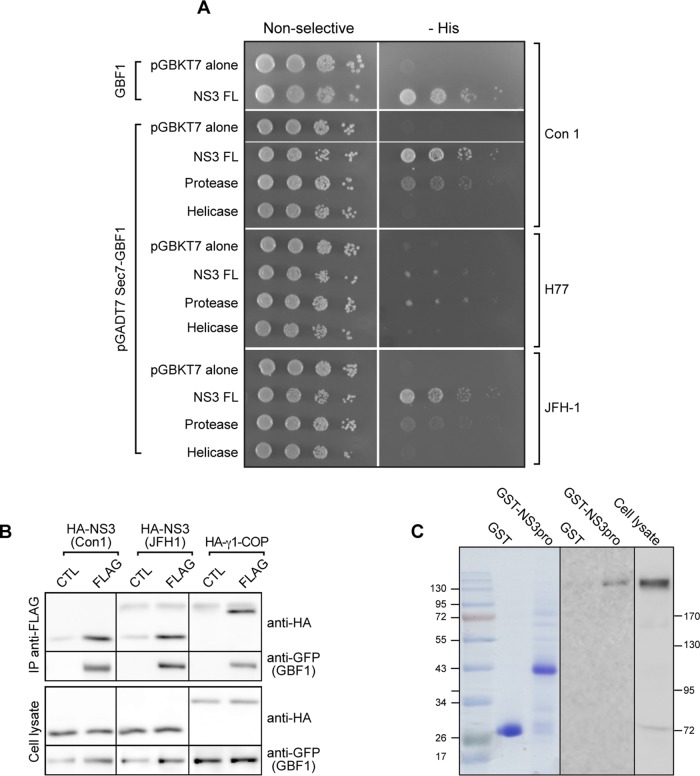
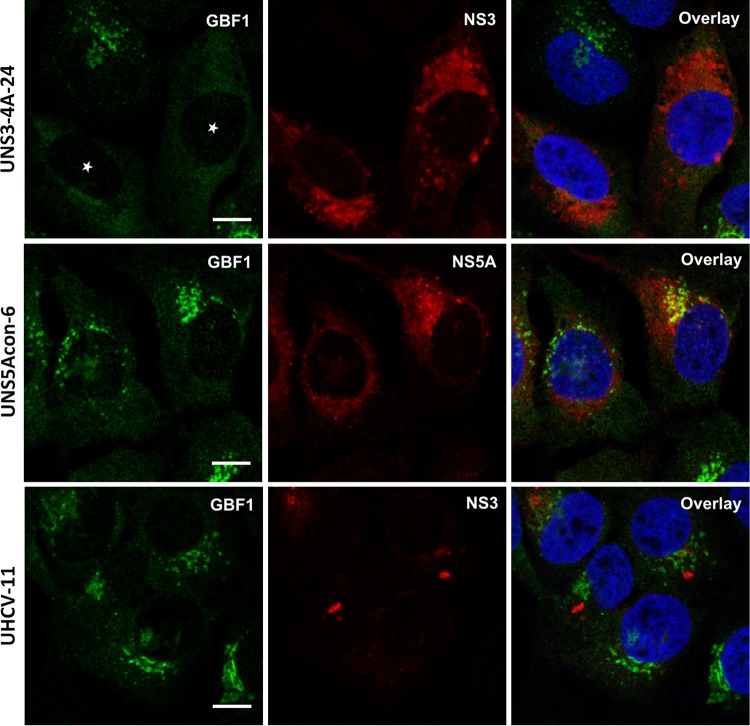
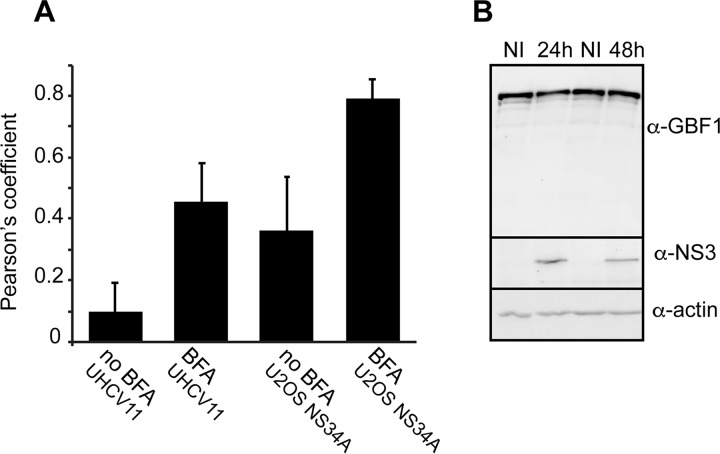
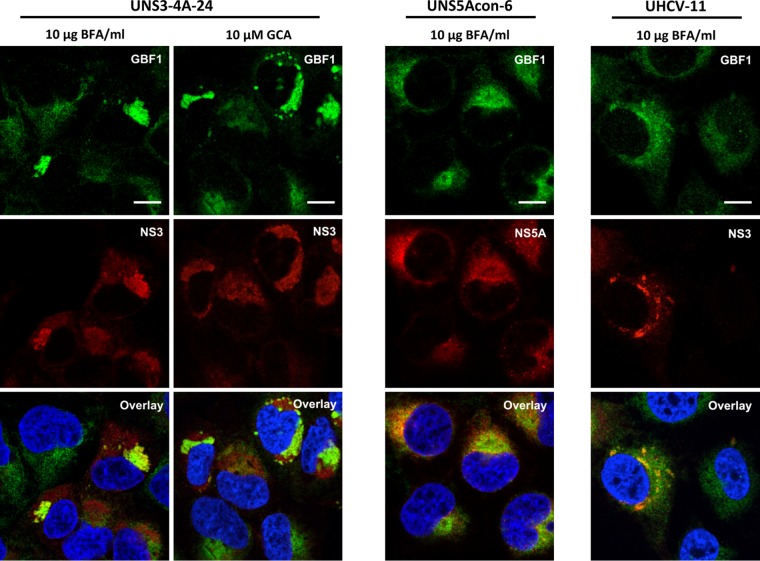
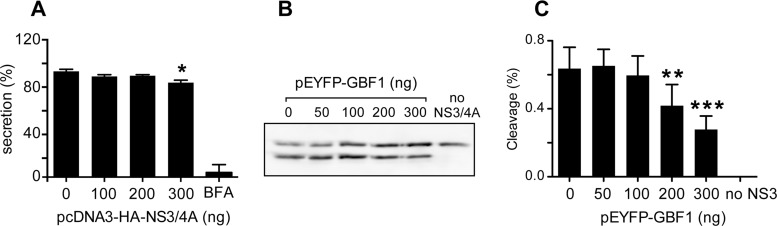
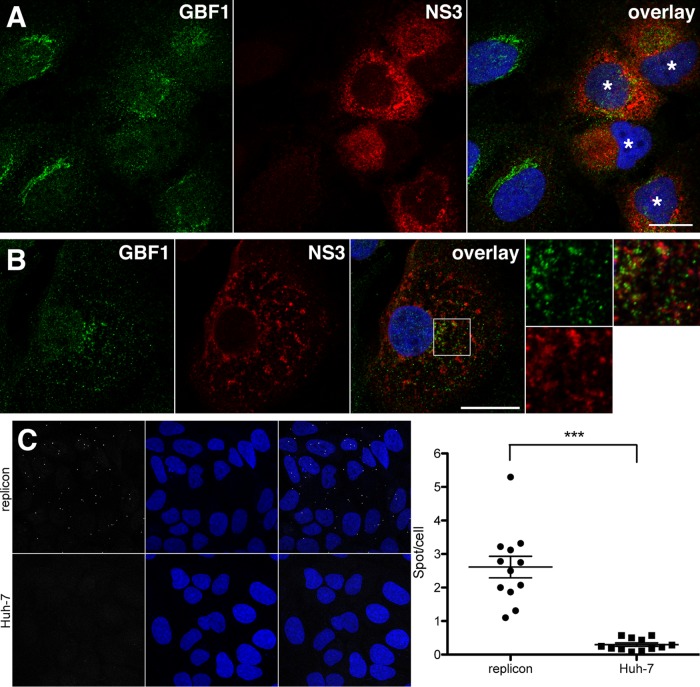
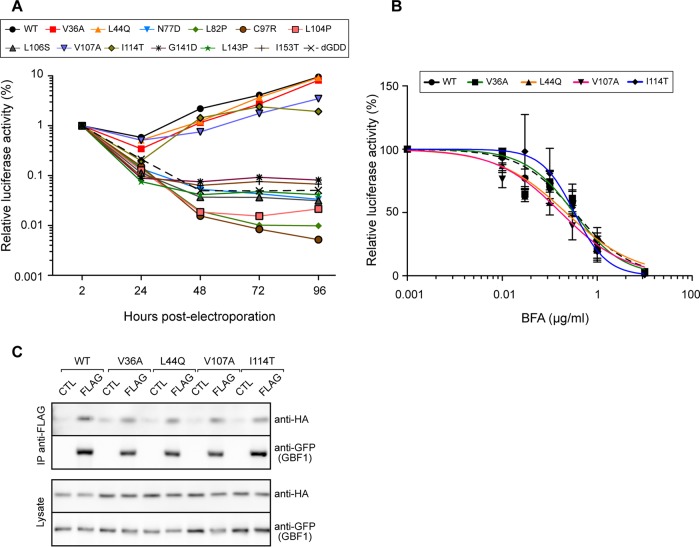
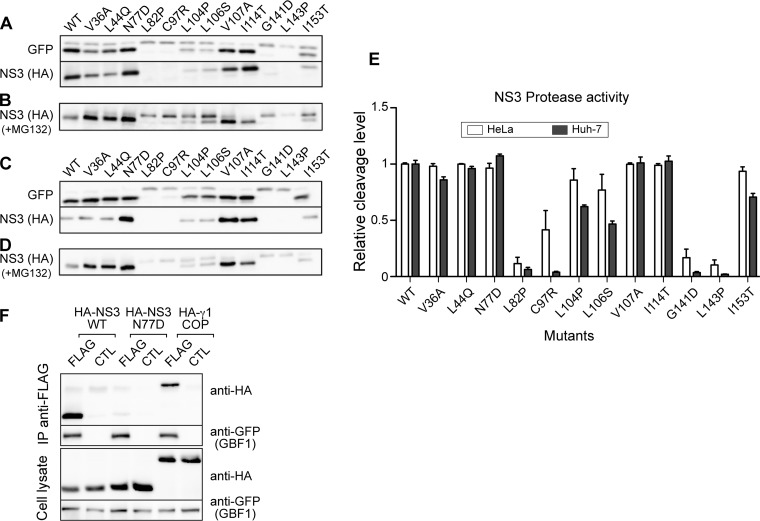
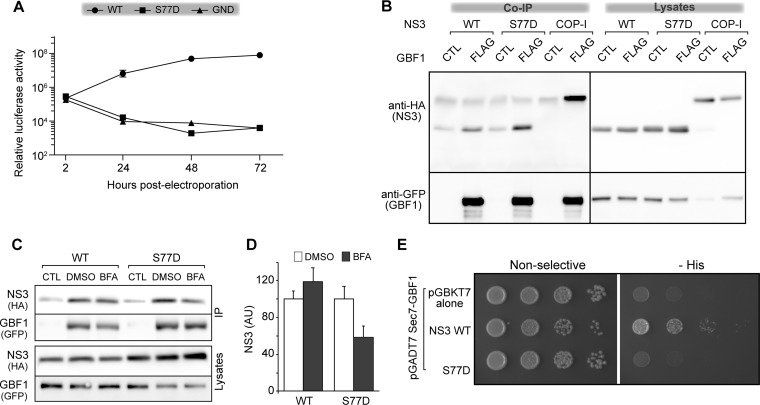
GBF1 has emerged as a host factor required for the genome replication of RNA viruses of different families. During the hepatitis C virus (HCV) life cycle, GBF1 performs a critical function at the onset of genome replication but is dispensable when the replication is established. To better understand how GBF1 regulates HCV infection, we have looked for interactions between GBF1 and HCV proteins. NS3 was found to interact with GBF1 in yeast two-hybrid, coimmunoprecipitation, and proximity ligation assays and to interfere with GBF1 function and alter GBF1 intracellular localization in cells expressing NS3. The interaction was mapped to the Sec7 domain of GBF1 and the protease domain of NS3. A reverse yeast two-hybrid screen to identify mutations altering NS3-GBF1 interaction yielded an NS3 mutant (N77D, Con1 strain) that is nonreplicative despite conserved protease activity and does not interact with GBF1. The mutated residue is exposed at the surface of NS3, suggesting it is part of the domain of NS3 that interacts with GBF1. The corresponding mutation in strain JFH-1 (S77D) produces a similar phenotype. Our results provide evidence for an interaction between NS3 and GBF1 and suggest that an alteration of this interaction is detrimental to HCV genome replication.IMPORTANCE Single-stranded, positive-sense RNA viruses rely to a significant extent on host factors to achieve the replication of their genome. GBF1 is such a cellular protein that is required for the replication of several RNA viruses, but its mechanism of action during viral infections is not yet defined. In this study, we investigated potential interactions that GBF1 might engage in with proteins of HCV, a GBF1-dependent virus. We found that GBF1 interacts with NS3, a nonstructural protein involved in HCV genome replication, and our results suggest that this interaction is important for GBF1 function during HCV replication. Interestingly, GBF1 interaction with HCV appears different from its interaction with enteroviruses, another group of GBF1-dependent RNA viruses, in keeping with the fact that HCV and enteroviruses use different functions of GBF1.
Keywords: ADP ribosylation factor; GBF1; NS3; hepatitis C virus; viral replication.
Copyright © 2019 American Society for Microbiology.
Figures

Similar articles
-
Hepatitis C Viral Replication Complex.Viruses. 2021 Mar 22;13(3):520. doi: 10.3390/v13030520. Viruses. 2021. PMID: 33809897 Free PMC article. Review.
-
Role of the Guanine Nucleotide Exchange Factor GBF1 in the Replication of RNA Viruses.Viruses. 2020 Jun 24;12(6):682. doi: 10.3390/v12060682. Viruses. 2020. PMID: 32599855 Free PMC article. Review.
-
Investigation of the role of GBF1 in the replication of positive-sense single-stranded RNA viruses.J Gen Virol. 2018 Aug;99(8):1086-1096. doi: 10.1099/jgv.0.001099. Epub 2018 Jun 20. J Gen Virol. 2018. PMID: 29923822
-
NS3 from Hepatitis C Virus Strain JFH-1 Is an Unusually Robust Helicase That Is Primed To Bind and Unwind Viral RNA.J Virol. 2017 Dec 14;92(1):e01253-17. doi: 10.1128/JVI.01253-17. Print 2018 Jan 1. J Virol. 2017. PMID: 29070684 Free PMC article.
-
A Redundant Mechanism of Recruitment Underlies the Remarkable Plasticity of the Requirement of Poliovirus Replication for the Cellular ArfGEF GBF1.J Virol. 2019 Oct 15;93(21):e00856-19. doi: 10.1128/JVI.00856-19. Print 2019 Nov 1. J Virol. 2019. PMID: 31375590 Free PMC article.
Cited by
-
A reporter cell line for the automated quantification of SARS-CoV-2 infection in living cells.Front Microbiol. 2022 Sep 29;13:1031204. doi: 10.3389/fmicb.2022.1031204. eCollection 2022. Front Microbiol. 2022. PMID: 36246297 Free PMC article.
-
The Guanine Nucleotide Exchange Factor GBF1 Participates in Rotavirus Replication.J Virol. 2019 Sep 12;93(19):e01062-19. doi: 10.1128/JVI.01062-19. Print 2019 Oct 1. J Virol. 2019. PMID: 31270230 Free PMC article.
-
Hepatitis C Viral Replication Complex.Viruses. 2021 Mar 22;13(3):520. doi: 10.3390/v13030520. Viruses. 2021. PMID: 33809897 Free PMC article. Review.
-
Viral use and subversion of membrane organization and trafficking.J Cell Sci. 2021 Mar 4;134(5):jcs252676. doi: 10.1242/jcs.252676. J Cell Sci. 2021. PMID: 33664154 Free PMC article. Review.
-
Role of the Guanine Nucleotide Exchange Factor GBF1 in the Replication of RNA Viruses.Viruses. 2020 Jun 24;12(6):682. doi: 10.3390/v12060682. Viruses. 2020. PMID: 32599855 Free PMC article. Review.
References
-
- Romero-Brey I, Merz A, Chiramel A, Lee JY, Chlanda P, Haselman U, Santarella-Mellwig R, Habermann A, Hoppe S, Kallis S, Walther P, Antony C, Krijnse-Locker J, Bartenschlager R. 2012. Three-dimensional architecture and biogenesis of membrane structures associated with hepatitis C virus replication. PLoS Pathog 8:e1003056. doi:10.1371/journal.ppat.1003056. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
