

HDX-MS reveals dysregulated checkpoints that compromise discrimination against self RNA during RIG-I mediated autoimmunity
- PMID: 30560918
- PMCID: PMC6299088
- DOI: 10.1038/s41467-018-07780-z
HDX-MS reveals dysregulated checkpoints that compromise discrimination against self RNA during RIG-I mediated autoimmunity
Abstract
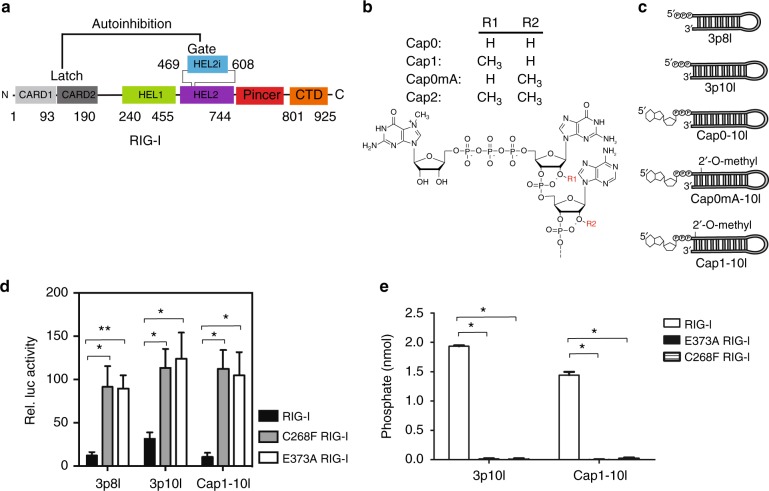
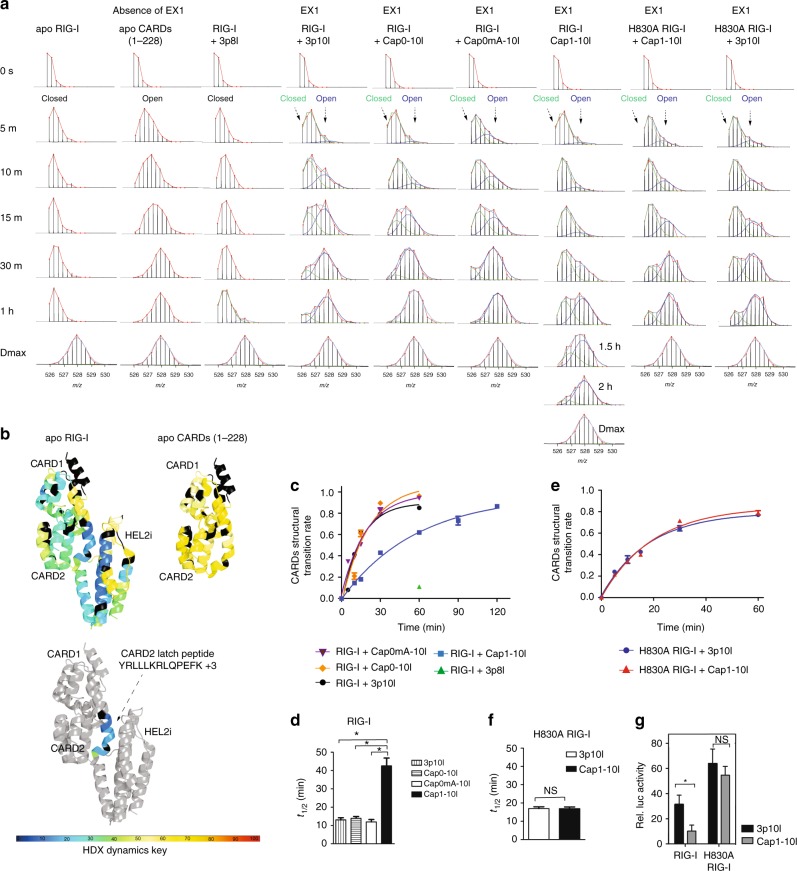
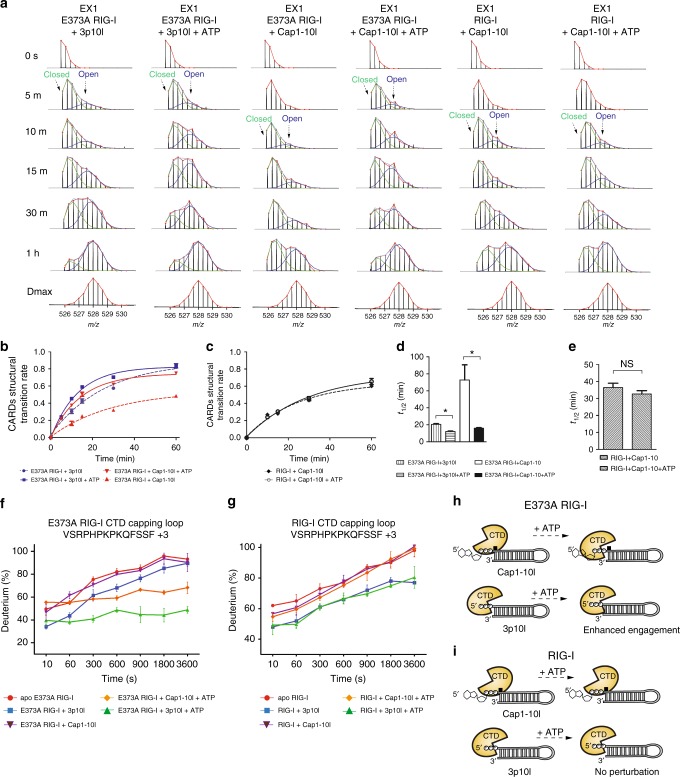
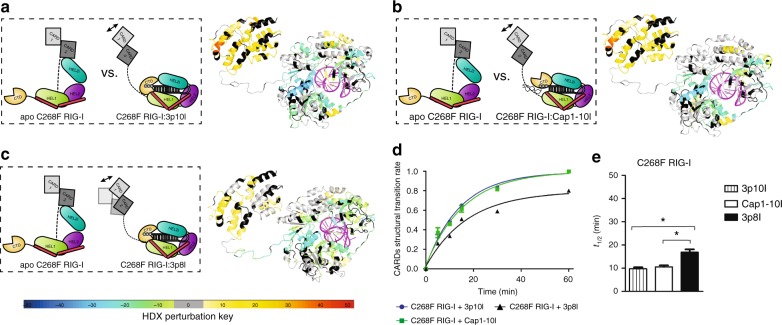
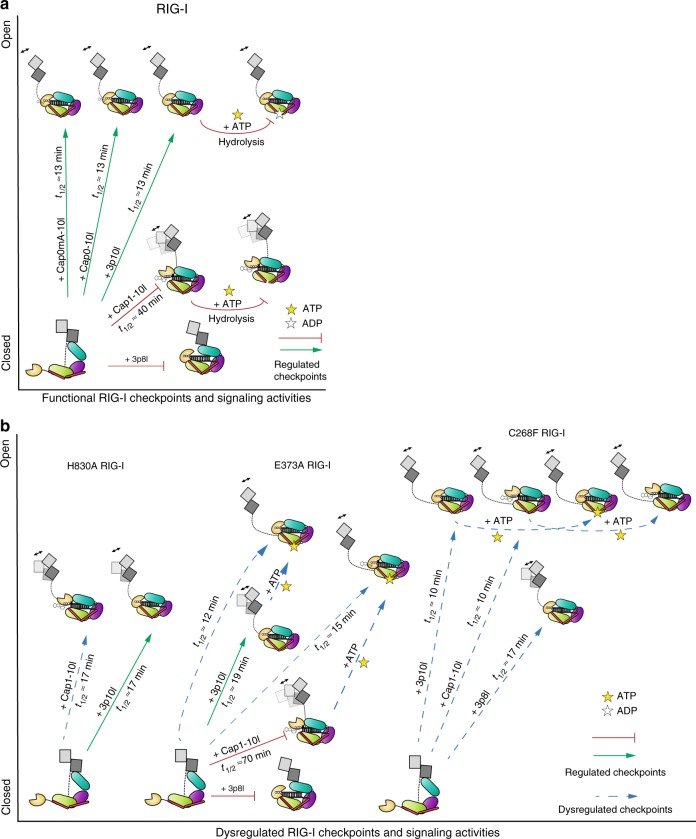
Retinoic acid inducible gene-I (RIG-I) ensures immune surveillance of viral RNAs bearing a 5'-triphosphate (5'ppp) moiety. Mutations in RIG-I (C268F and E373A) lead to impaired ATPase activity, thereby driving hyperactive signaling associated with autoimmune diseases. Here we report, using hydrogen/deuterium exchange, mechanistic models for dysregulated RIG-I proofreading that ultimately result in the improper recognition of cellular RNAs bearing 7-methylguanosine and N1-2'-O-methylation (Cap1) on the 5' end. Cap1-RNA compromises its ability to stabilize RIG-I helicase and blunts caspase activation and recruitment domains (CARD) partial opening by threefold. RIG-I H830A mutation restores Cap1-helicase engagement as well as CARDs partial opening event to a level comparable to that of 5'ppp. However, E373A RIG-I locks the receptor in an ATP-bound state, resulting in enhanced Cap1-helicase engagement and a sequential CARDs stimulation. C268F mutation renders a more tethered ring architecture and results in constitutive CARDs signaling in an ATP-independent manner.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Proofreading mechanisms of the innate immune receptor RIG-I: distinguishing self and viral RNA.Biochem Soc Trans. 2024 Jun 26;52(3):1131-1148. doi: 10.1042/BST20230724. Biochem Soc Trans. 2024. PMID: 38884803 Free PMC article. Review.
-
Structural basis for m7G recognition and 2'-O-methyl discrimination in capped RNAs by the innate immune receptor RIG-I.Proc Natl Acad Sci U S A. 2016 Jan 19;113(3):596-601. doi: 10.1073/pnas.1515152113. Epub 2016 Jan 5. Proc Natl Acad Sci U S A. 2016. PMID: 26733676 Free PMC article.
-
Kinetic discrimination of self/non-self RNA by the ATPase activity of RIG-I and MDA5.BMC Biol. 2015 Jul 28;13:54. doi: 10.1186/s12915-015-0166-9. BMC Biol. 2015. PMID: 26215161 Free PMC article.
-
RIG-I Uses an ATPase-Powered Translocation-Throttling Mechanism for Kinetic Proofreading of RNAs and Oligomerization.Mol Cell. 2018 Oct 18;72(2):355-368.e4. doi: 10.1016/j.molcel.2018.08.021. Epub 2018 Sep 27. Mol Cell. 2018. PMID: 30270105 Free PMC article.
-
Structures of RIG-I-Like Receptors and Insights into Viral RNA Sensing.Adv Exp Med Biol. 2019;1172:157-188. doi: 10.1007/978-981-13-9367-9_8. Adv Exp Med Biol. 2019. PMID: 31628656 Review.
Cited by
-
Proteomic Investigation of Immune Checkpoints and Some of Their Inhibitors.Int J Mol Sci. 2024 Aug 27;25(17):9276. doi: 10.3390/ijms25179276. Int J Mol Sci. 2024. PMID: 39273224 Free PMC article. Review.
-
N6-methyladenosine modification enables viral RNA to escape recognition by RNA sensor RIG-I.Nat Microbiol. 2020 Apr;5(4):584-598. doi: 10.1038/s41564-019-0653-9. Epub 2020 Feb 3. Nat Microbiol. 2020. PMID: 32015498 Free PMC article.
-
The intrinsically disordered CARDs-Helicase linker in RIG-I is a molecular gate for RNA proofreading.EMBO J. 2022 May 16;41(10):e109782. doi: 10.15252/embj.2021109782. Epub 2022 Apr 19. EMBO J. 2022. PMID: 35437807 Free PMC article.
-
Activation of cytosolic RNA sensors by endogenous ligands: roles in disease pathogenesis.Front Immunol. 2023 May 24;14:1092790. doi: 10.3389/fimmu.2023.1092790. eCollection 2023. Front Immunol. 2023. PMID: 37292201 Free PMC article. Review.
-
Proofreading mechanisms of the innate immune receptor RIG-I: distinguishing self and viral RNA.Biochem Soc Trans. 2024 Jun 26;52(3):1131-1148. doi: 10.1042/BST20230724. Biochem Soc Trans. 2024. PMID: 38884803 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
