

Endogenous DNA Double-Strand Breaks during DNA Transactions: Emerging Insights and Methods for Genome-Wide Profiling
- PMID: 30558210
- PMCID: PMC6316733
- DOI: 10.3390/genes9120632
Endogenous DNA Double-Strand Breaks during DNA Transactions: Emerging Insights and Methods for Genome-Wide Profiling
Abstract
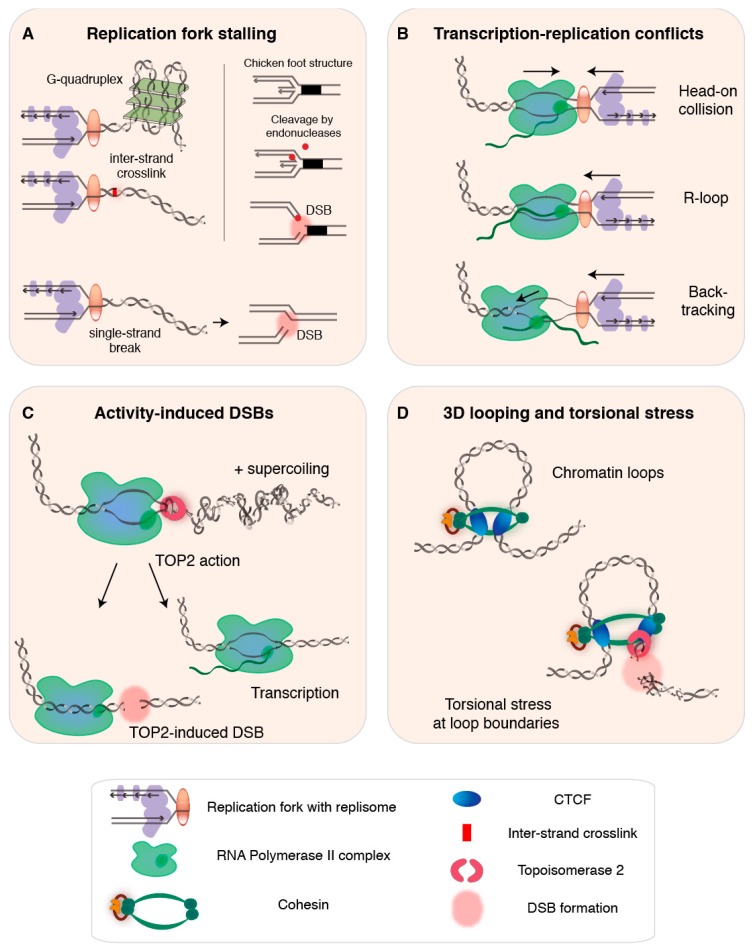
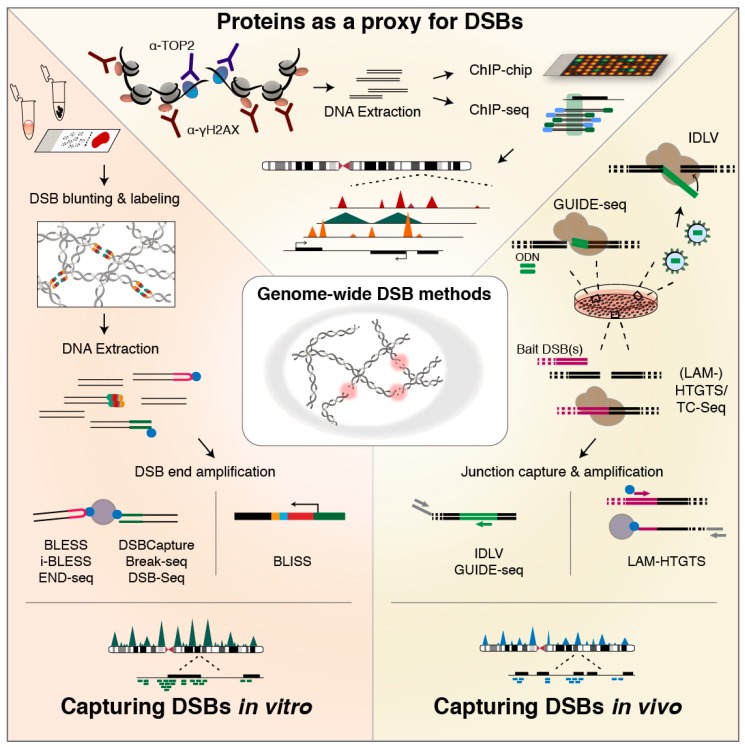
DNA double-strand breaks (DSBs) jeopardize genome integrity and can-when repaired unfaithfully-give rise to structural rearrangements associated with cancer. Exogenous agents such as ionizing radiation or chemotherapy can invoke DSBs, but a vast amount of breakage arises during vital endogenous DNA transactions, such as replication and transcription. Additionally, chromatin looping involved in 3D genome organization and gene regulation is increasingly recognized as a possible contributor to DSB events. In this review, we first discuss insights into the mechanisms of endogenous DSB formation, showcasing the trade-off between essential DNA transactions and the intrinsic challenges that these processes impose on genomic integrity. In the second part, we highlight emerging methods for genome-wide profiling of DSBs, and discuss future directions of research that will help advance our understanding of genome-wide DSB formation and repair.
Keywords: 3D genome structure; DNA double-strand breaks; cancer; chromatin looping; genome-wide methods; rearrangements; replication; topoisomerases; transcription.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
Ionizing radiation and genetic risks. XVII. Formation mechanisms underlying naturally occurring DNA deletions in the human genome and their potential relevance for bridging the gap between induced DNA double-strand breaks and deletions in irradiated germ cells.Mutat Res. 2013 Oct-Dec;753(2):114-130. doi: 10.1016/j.mrrev.2013.07.003. Epub 2013 Aug 12. Mutat Res. 2013. PMID: 23948232 Review.
-
Processing-Challenges Generated by Clusters of DNA Double-Strand Breaks Underpin Increased Effectiveness of High-LET Radiation and Chromothripsis.Adv Exp Med Biol. 2018;1044:149-168. doi: 10.1007/978-981-13-0593-1_10. Adv Exp Med Biol. 2018. PMID: 29956296 Review.
-
The Determinant of DNA Repair Pathway Choices in Ionising Radiation-Induced DNA Double-Strand Breaks.Biomed Res Int. 2020 Aug 25;2020:4834965. doi: 10.1155/2020/4834965. eCollection 2020. Biomed Res Int. 2020. PMID: 32908893 Free PMC article. Review.
-
Genomic rearrangements induced by unscheduled DNA double strand breaks in somatic mammalian cells.FEBS J. 2017 Aug;284(15):2324-2344. doi: 10.1111/febs.14053. Epub 2017 Mar 22. FEBS J. 2017. PMID: 28244221 Review.
-
Essential Role for an Isoform of Escherichia coli Translation Initiation Factor IF2 in Repair of Two-Ended DNA Double-Strand Breaks.J Bacteriol. 2022 Apr 19;204(4):e0057121. doi: 10.1128/jb.00571-21. Epub 2022 Mar 28. J Bacteriol. 2022. PMID: 35343794 Free PMC article.
Cited by
-
Non-homologous End Joining-Mediated Insertional Mutagenesis Reveals a Novel Target for Enhancing Fatty Alcohols Production in Yarrowia lipolytica.Front Microbiol. 2022 Apr 25;13:898884. doi: 10.3389/fmicb.2022.898884. eCollection 2022. Front Microbiol. 2022. PMID: 35547152 Free PMC article.
-
Broken by the Cut: A Journey into the Role of Topoisomerase II in DNA Fragility.Genes (Basel). 2019 Oct 12;10(10):791. doi: 10.3390/genes10100791. Genes (Basel). 2019. PMID: 31614754 Free PMC article. Review.
-
Somatic Copy Number Alterations in Human Cancers: An Analysis of Publicly Available Data From The Cancer Genome Atlas.Front Oncol. 2021 Jul 28;11:700568. doi: 10.3389/fonc.2021.700568. eCollection 2021. Front Oncol. 2021. PMID: 34395272 Free PMC article.
-
Activation of Oncogenic Super-Enhancers Is Coupled with DNA Repair by RAD51.Cell Rep. 2019 Oct 15;29(3):560-572.e4. doi: 10.1016/j.celrep.2019.09.001. Cell Rep. 2019. PMID: 31618627 Free PMC article.
-
Transcription is a major driving force for plastid genome instability in Arabidopsis.PLoS One. 2019 Apr 3;14(4):e0214552. doi: 10.1371/journal.pone.0214552. eCollection 2019. PLoS One. 2019. PMID: 30943245 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
