

TGNap1 is required for microtubule-dependent homeostasis of a subpopulation of the plant trans-Golgi network
- PMID: 30552321
- PMCID: PMC6294250
- DOI: 10.1038/s41467-018-07662-4
TGNap1 is required for microtubule-dependent homeostasis of a subpopulation of the plant trans-Golgi network
Abstract
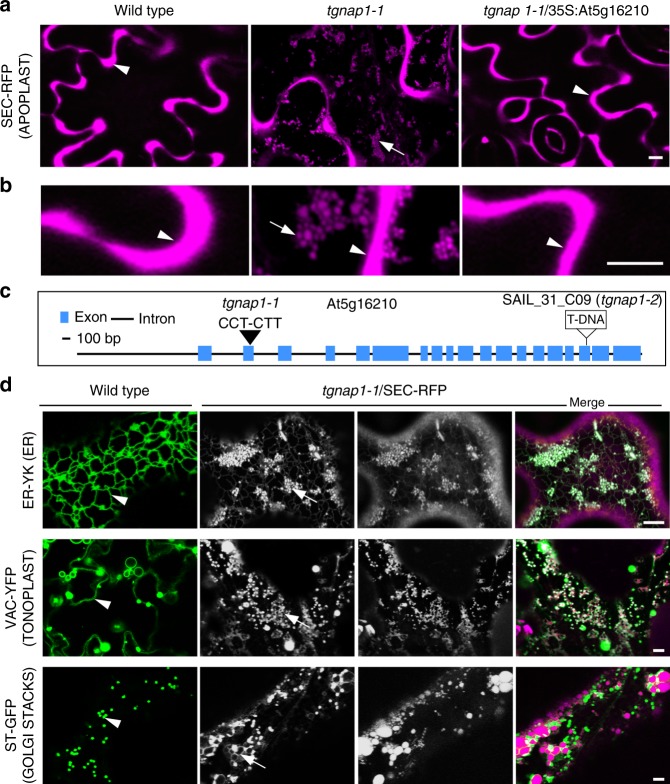
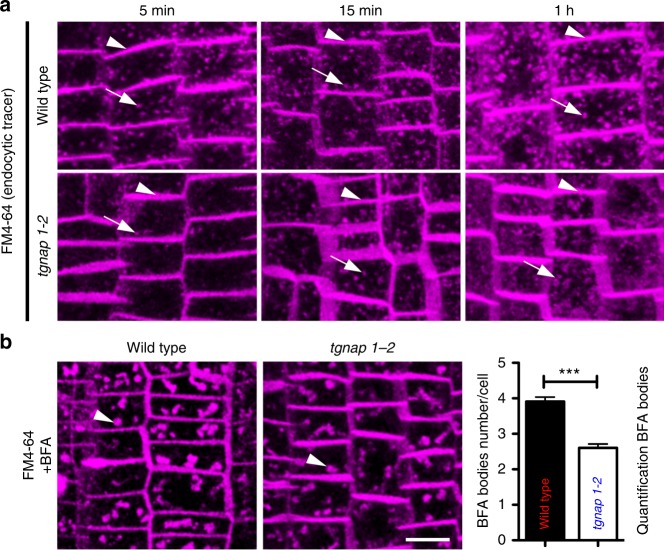
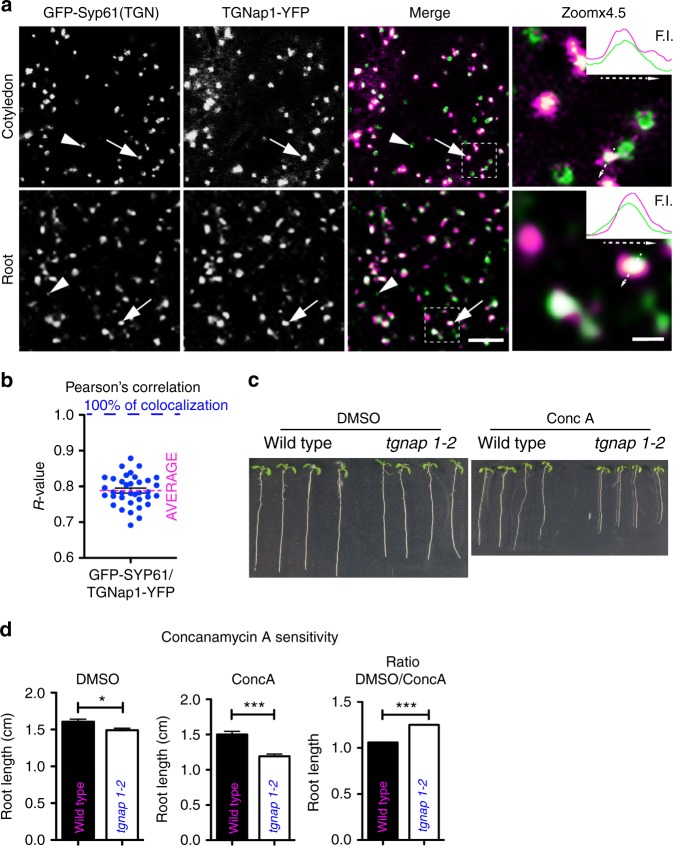
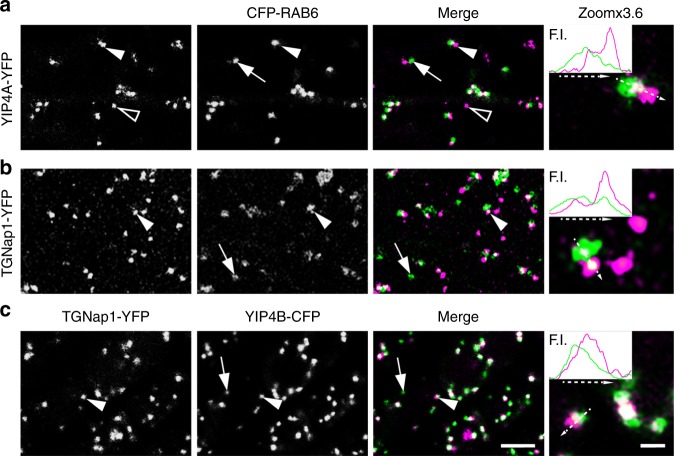
Defining convergent and divergent mechanisms underlying the biogenesis and function of endomembrane organelles is fundamentally important in cell biology. In all eukaryotes, the Trans-Golgi Network (TGN) is the hub where the exocytic and endocytic pathways converge. To gain knowledge in the mechanisms underlying TGN biogenesis and function, we characterized TGNap1, a protein encoded by a plant gene of unknown function conserved with metazoans. We demonstrate that TGNap1 is a TGN protein required for the homeostasis of biosynthetic and endocytic traffic pathways. We also show that TGNap1 binds Rab6, YIP4 and microtubules. Finally, we establish that TGNap1 contributes to microtubule-dependent biogenesis, tracking and function of a TGN subset, likely through interaction with Rab6 and YIP4. Our results identify an important trafficking determinant at the plant TGN and reveal an unexpected reliance of post-Golgi traffic homeostasis and organelle biogenesis on microtubules in plants.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Linking secretion and cytoskeleton in immunity- a case for Arabidopsis TGNap1.Bioessays. 2024 Nov;46(11):e2400150. doi: 10.1002/bies.202400150. Epub 2024 Sep 20. Bioessays. 2024. PMID: 39302180
-
Defense against phytopathogens relies on efficient antimicrobial protein secretion mediated by the microtubule-binding protein TGNap1.Nat Commun. 2023 Oct 11;14(1):6357. doi: 10.1038/s41467-023-41807-4. Nat Commun. 2023. PMID: 37821453 Free PMC article.
-
A Golgi-Released Subpopulation of the Trans-Golgi Network Mediates Protein Secretion in Arabidopsis.Plant Physiol. 2019 Feb;179(2):519-532. doi: 10.1104/pp.18.01228. Epub 2018 Dec 13. Plant Physiol. 2019. PMID: 30545905 Free PMC article.
-
Arabidopsis TRAPPII is functionally linked to Rab-A, but not Rab-D in polar protein trafficking in trans-Golgi network.Plant Signal Behav. 2011 Nov;6(11):1679-83. doi: 10.4161/psb.6.11.17915. Epub 2011 Nov 1. Plant Signal Behav. 2011. PMID: 22067991 Free PMC article.
-
RAB GTPases and SNAREs at the trans-Golgi network in plants.J Plant Res. 2022 May;135(3):389-403. doi: 10.1007/s10265-022-01392-x. Epub 2022 Apr 29. J Plant Res. 2022. PMID: 35488138 Free PMC article. Review.
Cited by
-
ESCRT components ISTL1 andLIP5 are required for tapetal function and pollen viability.Plant Cell. 2021 Aug 31;33(8):2850-2868. doi: 10.1093/plcell/koab132. Plant Cell. 2021. PMID: 34125207 Free PMC article.
-
MTV proteins unveil ER- and microtubule-associated compartments in the plant vacuolar trafficking pathway.Proc Natl Acad Sci U S A. 2020 May 5;117(18):9884-9895. doi: 10.1073/pnas.1919820117. Epub 2020 Apr 22. Proc Natl Acad Sci U S A. 2020. PMID: 32321832 Free PMC article.
-
Fifteen compelling open questions in plant cell biology.Plant Cell. 2022 Jan 20;34(1):72-102. doi: 10.1093/plcell/koab225. Plant Cell. 2022. PMID: 34529074 Free PMC article. Review.
-
Actin and Microtubules Differently Contribute to Vacuolar Targeting Specificity during the Export from the ER.Membranes (Basel). 2021 Apr 20;11(4):299. doi: 10.3390/membranes11040299. Membranes (Basel). 2021. PMID: 33924184 Free PMC article.
-
Cargo sorting zones in the trans-Golgi network visualized by super-resolution confocal live imaging microscopy in plants.Nat Commun. 2021 Mar 26;12(1):1901. doi: 10.1038/s41467-021-22267-0. Nat Commun. 2021. PMID: 33772008 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
