

Genetic analysis reveals functions of atypical polyubiquitin chains
- PMID: 30547882
- PMCID: PMC6305200
- DOI: 10.7554/eLife.42955
Genetic analysis reveals functions of atypical polyubiquitin chains
Abstract
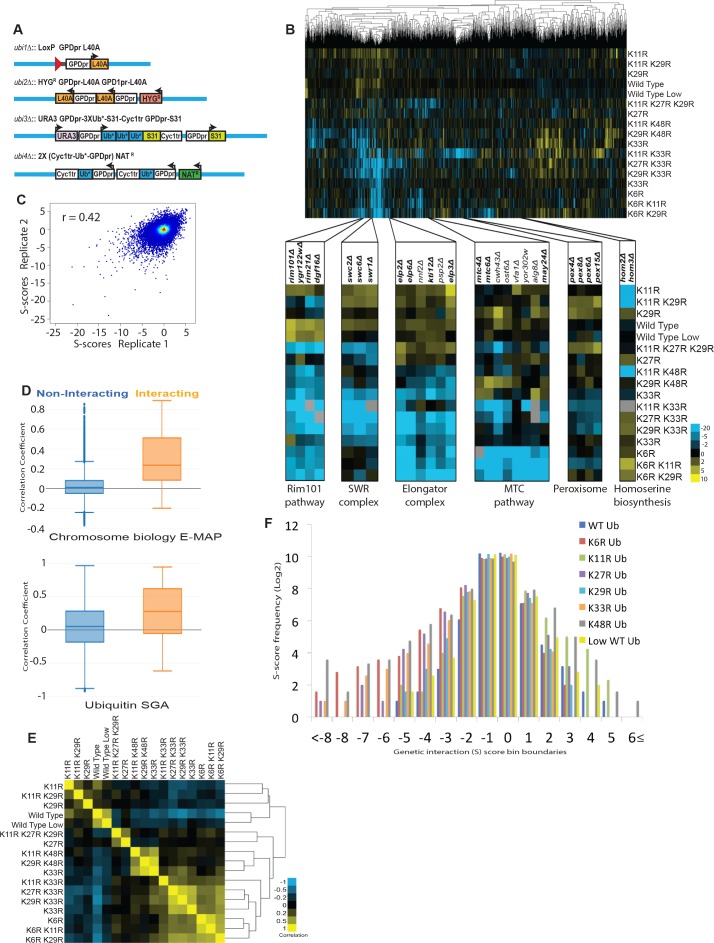
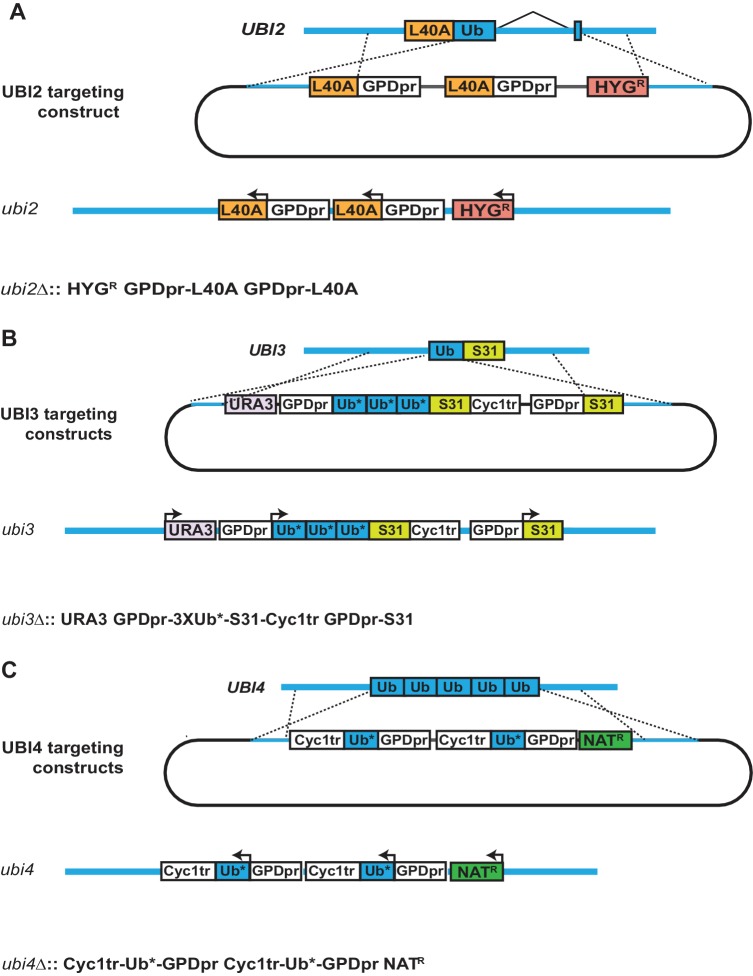
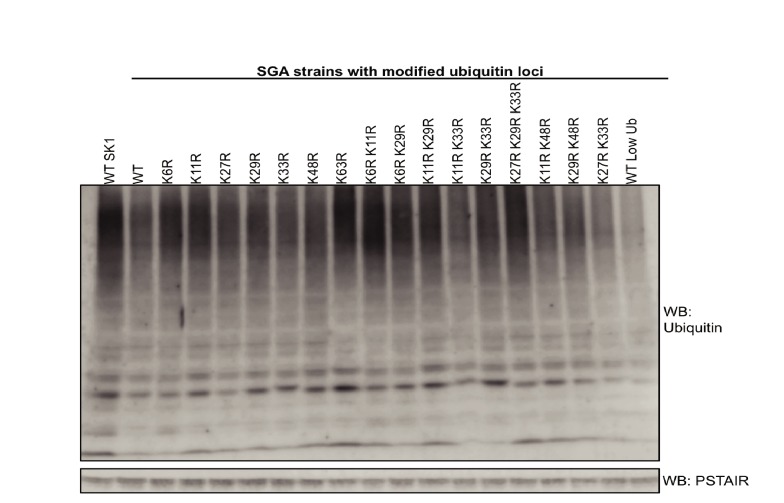
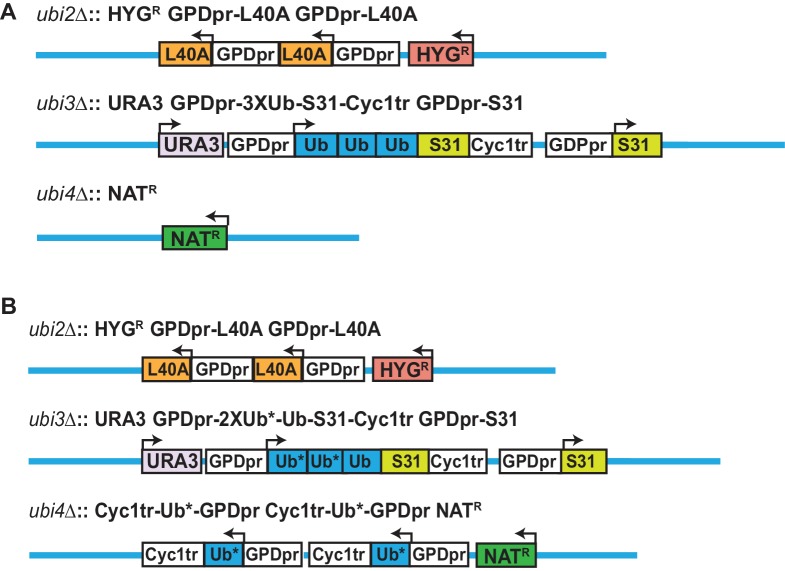
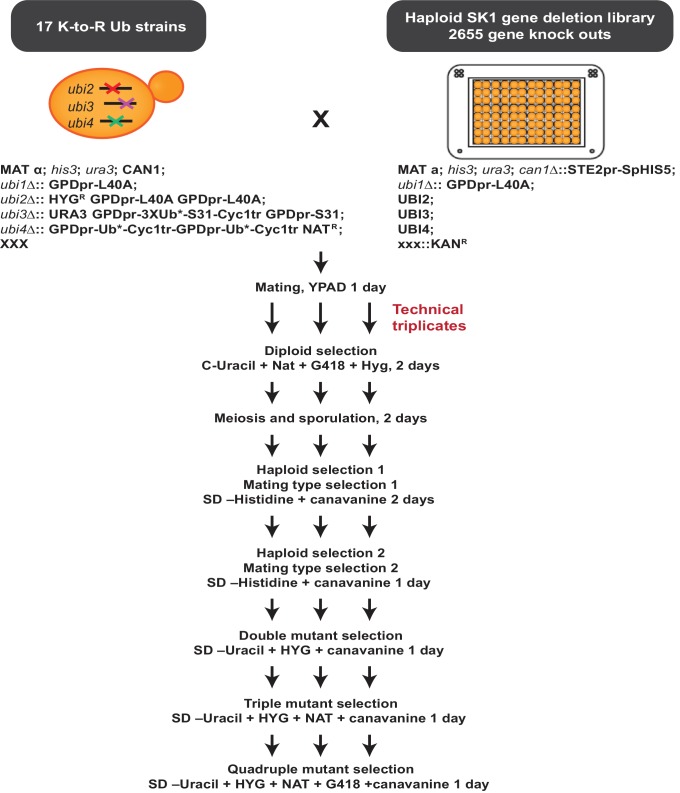
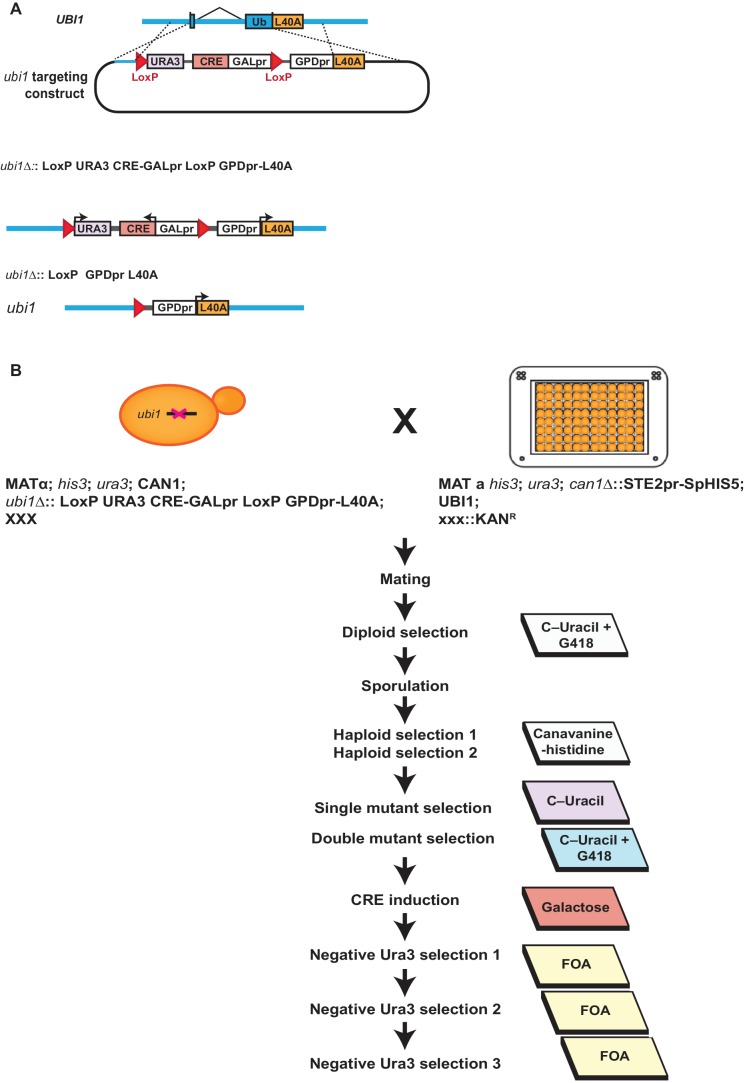
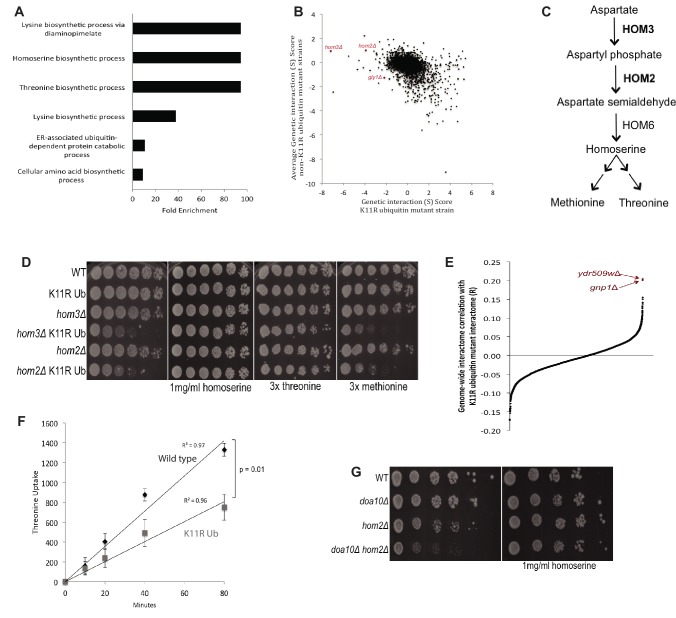
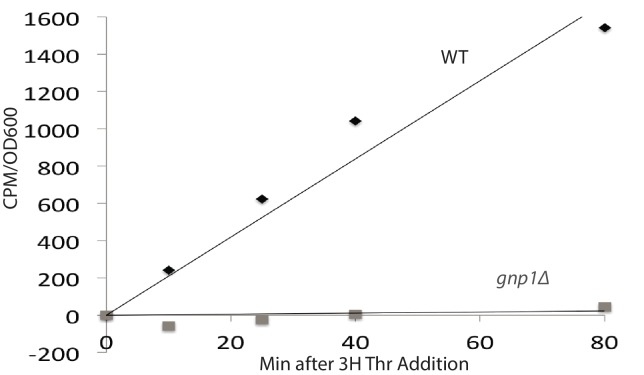
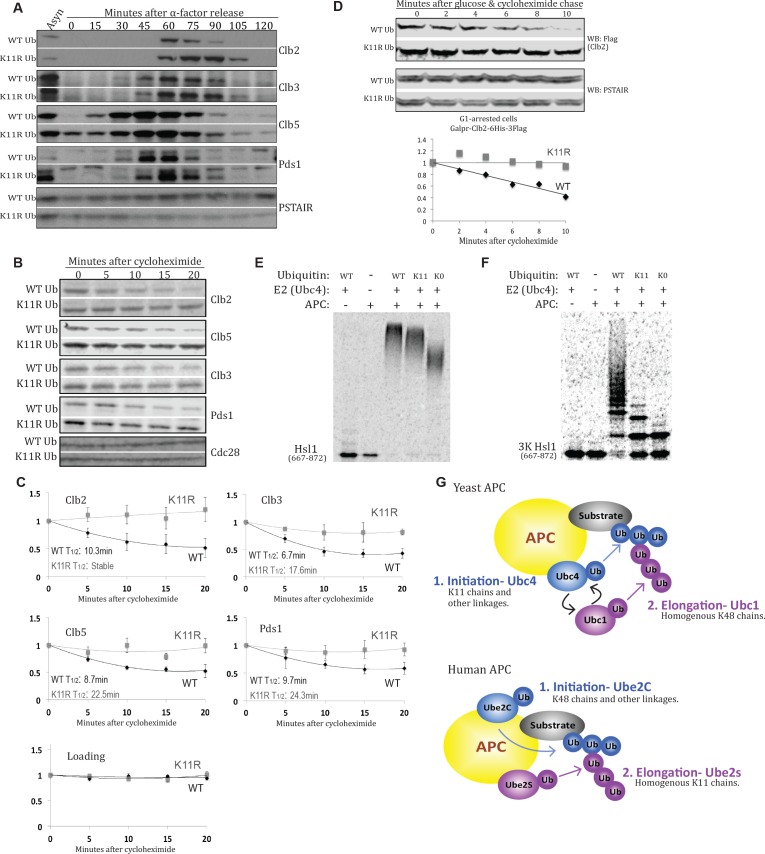
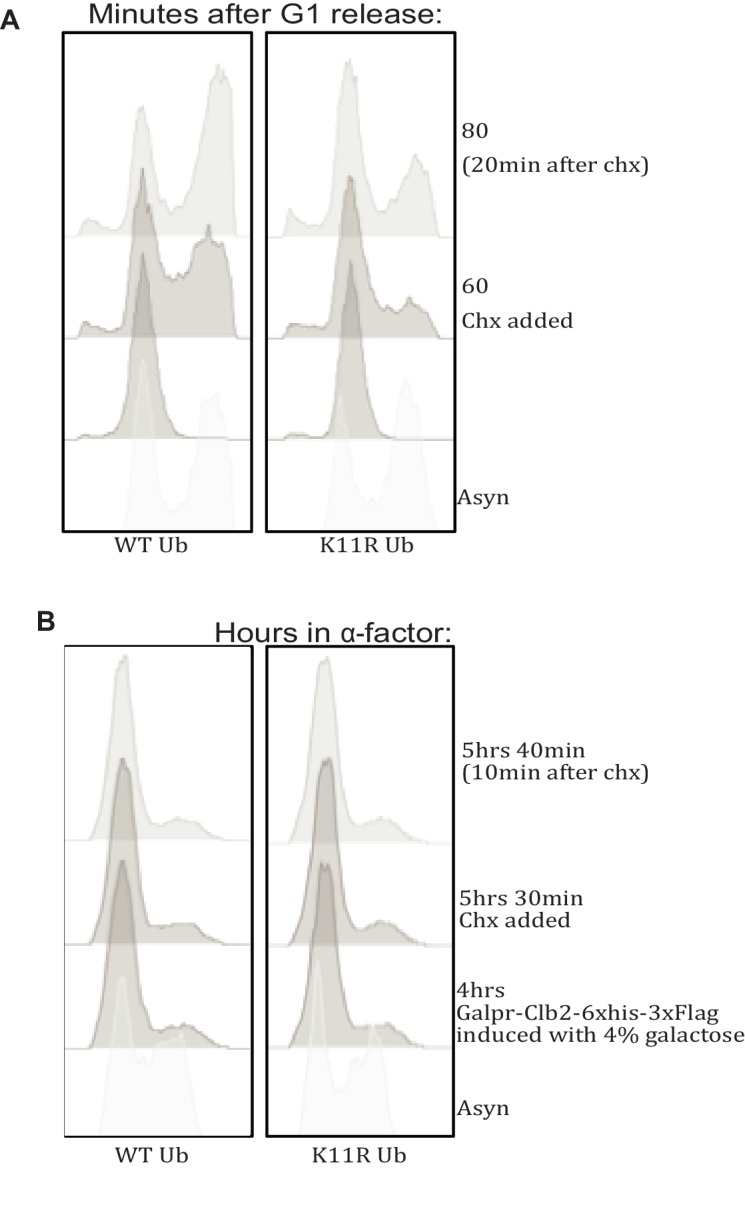
Although polyubiquitin chains linked through all lysines of ubiquitin exist, specific functions are well-established only for lysine-48 and lysine-63 linkages in Saccharomyces cerevisiae. To uncover pathways regulated by distinct linkages, genetic interactions between a gene deletion library and a panel of lysine-to-arginine ubiquitin mutants were systematically identified. The K11R mutant had strong genetic interactions with threonine biosynthetic genes. Consistently, we found that K11R mutants import threonine poorly. The K11R mutant also exhibited a strong genetic interaction with a subunit of the anaphase-promoting complex (APC), suggesting a role in cell cycle regulation. K11-linkages are important for vertebrate APC function, but this was not previously described in yeast. We show that the yeast APC also modifies substrates with K11-linkages in vitro, and that those chains contribute to normal APC-substrate turnover in vivo. This study reveals comprehensive genetic interactomes of polyubiquitin chains and characterizes the role of K11-chains in two biological pathways.
Keywords: APC; K11; S. cerevisiae; biochemistry; chemical biology; genetics; genomics; ubiquitin.
© 2018, Meza Gutierrez et al.
Conflict of interest statement
FM, AM, AD, HB, JJ, JX, MS, MN, RT, CP, LS, NK, DT No competing interests declared, DS is affiliated with Amgen Research. The author has no other competing interests to declare.
Figures

Similar articles
-
Efficient APC/C substrate degradation in cells undergoing mitotic exit depends on K11 ubiquitin linkages.Mol Biol Cell. 2015 Dec 1;26(24):4325-32. doi: 10.1091/mbc.E15-02-0102. Epub 2015 Oct 7. Mol Biol Cell. 2015. PMID: 26446837 Free PMC article.
-
A genetic approach to study polyubiquitination in Saccharomyces cerevisiae.Methods Enzymol. 2019;618:49-72. doi: 10.1016/bs.mie.2018.12.020. Epub 2019 Feb 21. Methods Enzymol. 2019. PMID: 30850062 Free PMC article.
-
Quantitative proteomics reveals the function of unconventional ubiquitin chains in proteasomal degradation.Cell. 2009 Apr 3;137(1):133-45. doi: 10.1016/j.cell.2009.01.041. Cell. 2009. PMID: 19345192 Free PMC article.
-
Assembly of k11-linked ubiquitin chains by the anaphase-promoting complex.Subcell Biochem. 2010;54:107-15. doi: 10.1007/978-1-4419-6676-6_9. Subcell Biochem. 2010. PMID: 21222277 Review.
-
K11-linked ubiquitin chains as novel regulators of cell division.Trends Cell Biol. 2011 Nov;21(11):656-63. doi: 10.1016/j.tcb.2011.08.008. Epub 2011 Oct 4. Trends Cell Biol. 2011. PMID: 21978762 Free PMC article. Review.
Cited by
-
Proteomic Analysis Reveals Sex-Specific Protein Degradation Targets in the Amygdala During Fear Memory Formation.Front Mol Neurosci. 2021 Sep 29;14:716284. doi: 10.3389/fnmol.2021.716284. eCollection 2021. Front Mol Neurosci. 2021. PMID: 34658783 Free PMC article.
-
APC/C: current understanding and future perspectives.F1000Res. 2019 May 23;8:F1000 Faculty Rev-725. doi: 10.12688/f1000research.18582.1. eCollection 2019. F1000Res. 2019. PMID: 31164978 Free PMC article. Review.
-
The diversity of linkage-specific polyubiquitin chains and their role in synaptic plasticity and memory formation.Neurobiol Learn Mem. 2020 Oct;174:107286. doi: 10.1016/j.nlm.2020.107286. Epub 2020 Aug 1. Neurobiol Learn Mem. 2020. PMID: 32745599 Free PMC article. Review.
-
Emerging Roles of Non-proteolytic Ubiquitination in Tumorigenesis.Front Cell Dev Biol. 2022 Jul 6;10:944460. doi: 10.3389/fcell.2022.944460. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35874839 Free PMC article. Review.
-
Mechanism of Lys6 poly-ubiquitin specificity by the L. pneumophila deubiquitinase LotA.Mol Cell. 2023 Jan 5;83(1):105-120.e5. doi: 10.1016/j.molcel.2022.11.022. Epub 2022 Dec 19. Mol Cell. 2023. PMID: 36538933 Free PMC article.
References
-
- Birsa N, Norkett R, Wauer T, Mevissen TE, Wu HC, Foltynie T, Bhatia K, Hirst WD, Komander D, Plun-Favreau H, Kittler JT. Lysine 27 ubiquitination of the mitochondrial transport protein Miro is dependent on serine 65 of the Parkin ubiquitin ligase. Journal of Biological Chemistry. 2014;289:14569–14582. doi: 10.1074/jbc.M114.563031. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
