

The reduced activity of PP-1α under redox stress condition is a consequence of GSH-mediated transient disulfide formation
- PMID: 30531830
- PMCID: PMC6286341
- DOI: 10.1038/s41598-018-36267-6
The reduced activity of PP-1α under redox stress condition is a consequence of GSH-mediated transient disulfide formation
Abstract
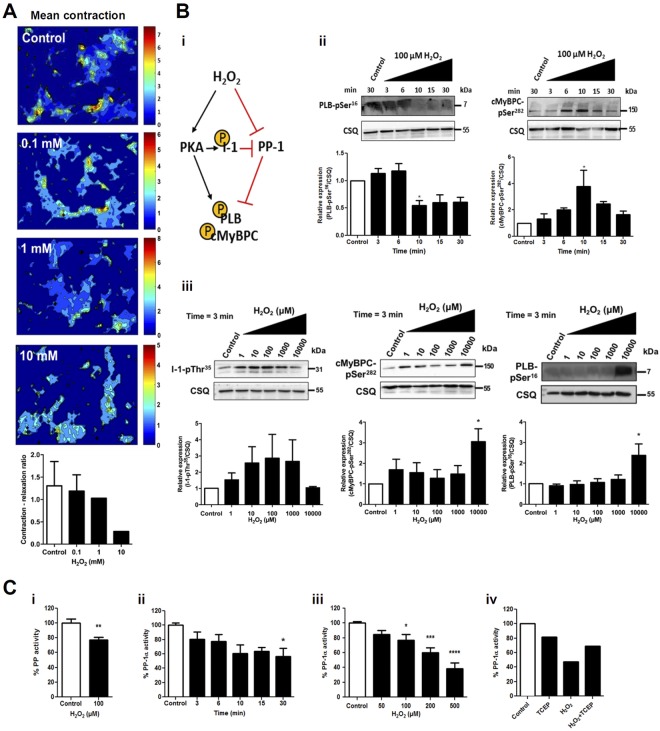
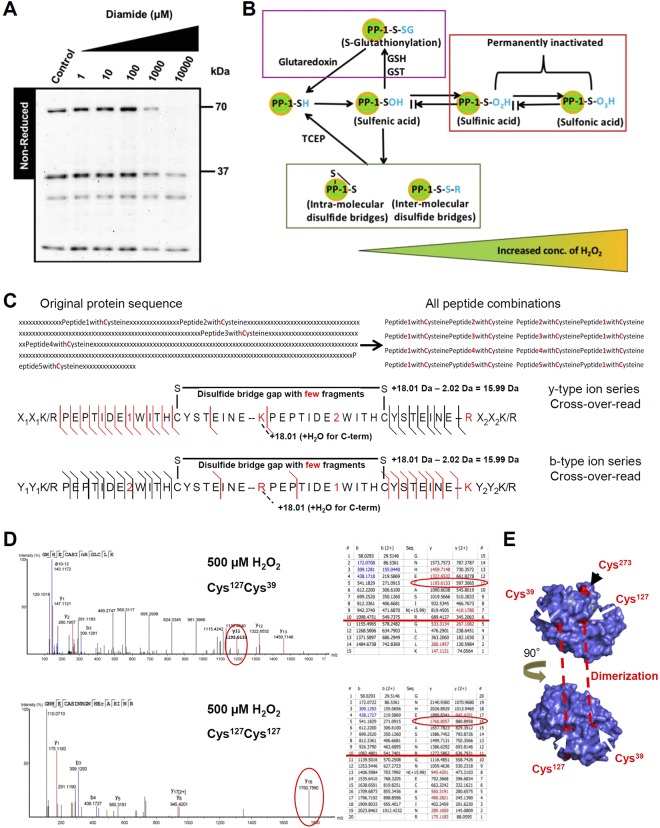
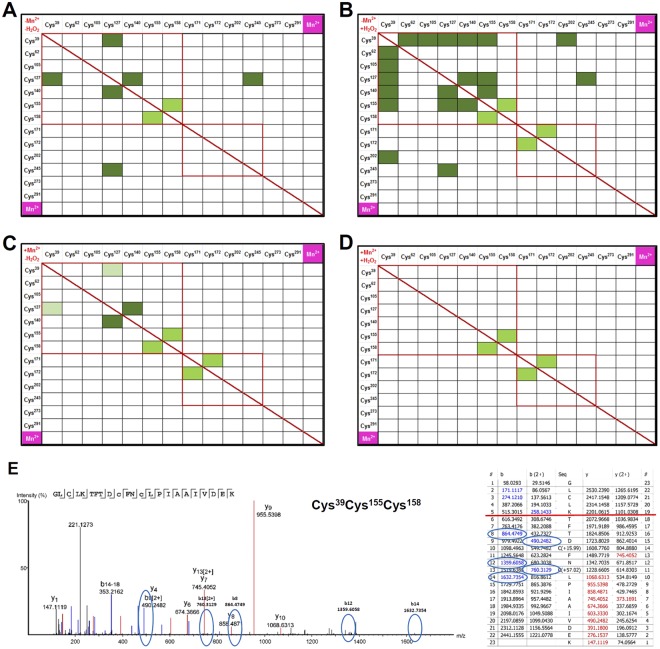
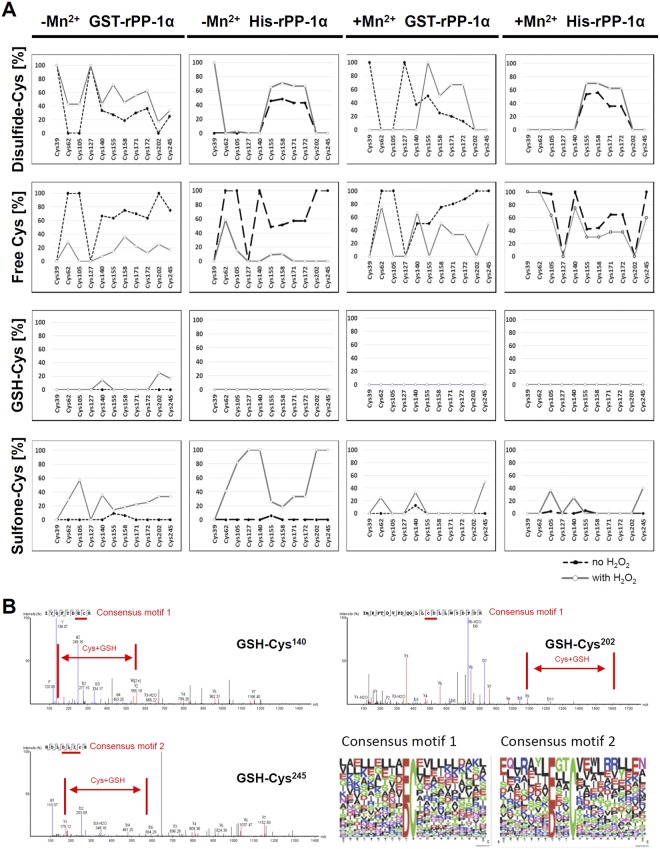
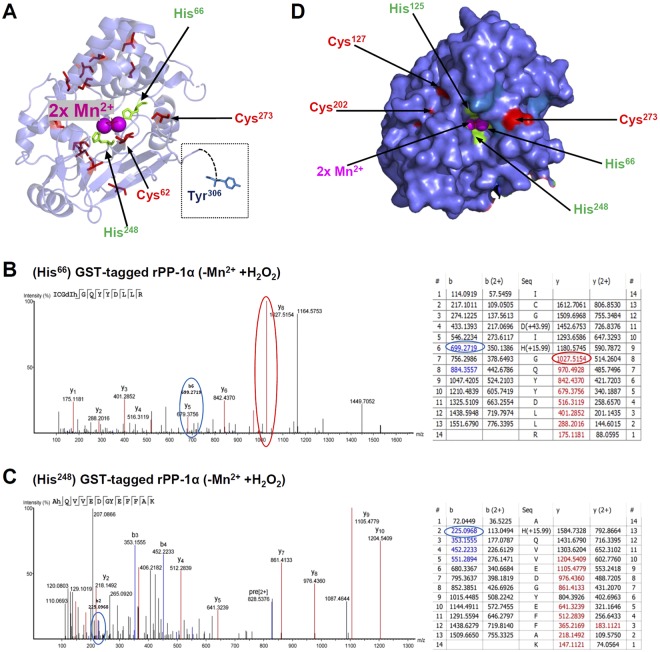
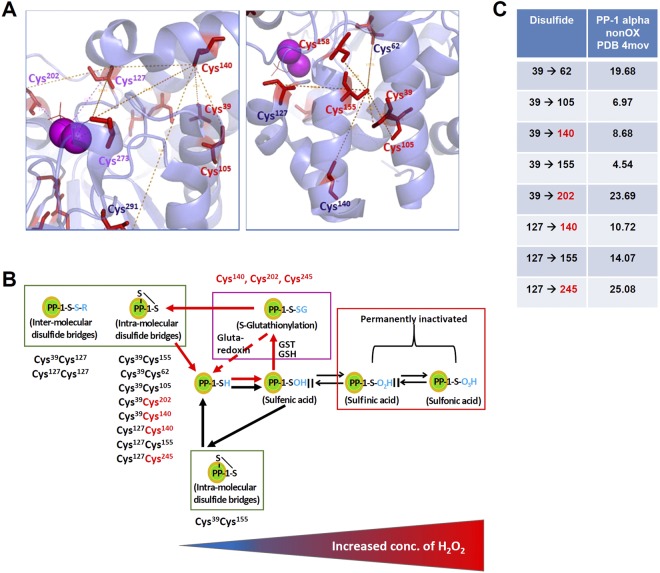
Heart failure is the most common cause of morbidity and hospitalization in the western civilization. Protein phosphatases play a key role in the basal cardiac contractility and in the responses to β-adrenergic stimulation with type-1 phosphatase (PP-1) being major contributor. We propose here that formation of transient disulfide bridges in PP-1α might play a leading role in oxidative stress response. First, we established an optimized workflow, the so-called "cross-over-read" search method, for the identification of disulfide-linked species using permutated databases. By applying this method, we demonstrate the formation of unexpected transient disulfides in PP-1α to shelter against over-oxidation. This protection mechanism strongly depends on the fast response in the presence of reduced glutathione. Our work points out that the dimerization of PP-1α involving Cys39 and Cys127 is presumably important for the protection of PP-1α active surface in the absence of a substrate. We finally give insight into the electron transport from the PP-1α catalytic core to the surface. Our data suggest that the formation of transient disulfides might be a general mechanism of proteins to escape from irreversible cysteine oxidation and to prevent their complete inactivation.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Reversible oxidation controls the activity and oligomeric state of the mammalian phosphoglycolate phosphatase AUM.Free Radic Biol Med. 2016 Aug;97:75-84. doi: 10.1016/j.freeradbiomed.2016.05.007. Epub 2016 May 11. Free Radic Biol Med. 2016. PMID: 27179418
-
Disulfide stress: a novel type of oxidative stress in acute pancreatitis.Free Radic Biol Med. 2014 May;70:265-77. doi: 10.1016/j.freeradbiomed.2014.01.009. Epub 2014 Jan 20. Free Radic Biol Med. 2014. PMID: 24456905
-
Formation and reduction of glutathione-protein mixed disulfides during oxidative stress. A study with isolated hepatocytes and menadione (2-methyl-1,4-naphthoquinone).Biochem Pharmacol. 1987 Apr 15;36(8):1313-20. doi: 10.1016/0006-2952(87)90087-6. Biochem Pharmacol. 1987. PMID: 3593416
-
Regulation of protein function by glutathionylation.Free Radic Res. 2005 Jun;39(6):573-80. doi: 10.1080/10715760500072172. Free Radic Res. 2005. PMID: 16036334 Review.
-
Role of ascorbate in oxidative protein folding.Biofactors. 2003;17(1-4):37-46. doi: 10.1002/biof.5520170105. Biofactors. 2003. PMID: 12897427 Review.
Cited by
-
Protein phosphatase-1 regulates the binding of filamin C to FILIP1 in cultured skeletal muscle cells under mechanical stress.Sci Rep. 2024 Nov 9;14(1):27348. doi: 10.1038/s41598-024-78953-8. Sci Rep. 2024. PMID: 39521905 Free PMC article.
-
Receptor-independent modulation of cAMP-dependent protein kinase and protein phosphatase signaling in cardiac myocytes by oxidizing agents.J Biol Chem. 2020 Nov 6;295(45):15342-15365. doi: 10.1074/jbc.RA120.014467. Epub 2020 Aug 31. J Biol Chem. 2020. PMID: 32868295 Free PMC article.
-
Mass Spectrometry-Based Redox and Protein Profiling of Failing Human Hearts.Int J Mol Sci. 2021 Feb 11;22(4):1787. doi: 10.3390/ijms22041787. Int J Mol Sci. 2021. PMID: 33670142 Free PMC article.
-
Regulation of Cardiac PKA Signaling by cAMP and Oxidants.Antioxidants (Basel). 2021 Apr 24;10(5):663. doi: 10.3390/antiox10050663. Antioxidants (Basel). 2021. PMID: 33923287 Free PMC article. Review.
References
-
- Griendling KK, FitzGerald GA. Oxidative stress and cardiovascular injury: Part I: basic mechanisms and in vivo monitoring of ROS. Circulation. 2003;108:1912–1916. doi: 10.1161/01.CIR.0000093660.86242.BB. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
