

Migration through a small pore disrupts inactive chromatin organization in neutrophil-like cells
- PMID: 30477489
- PMCID: PMC6257957
- DOI: 10.1186/s12915-018-0608-2
Migration through a small pore disrupts inactive chromatin organization in neutrophil-like cells
Abstract
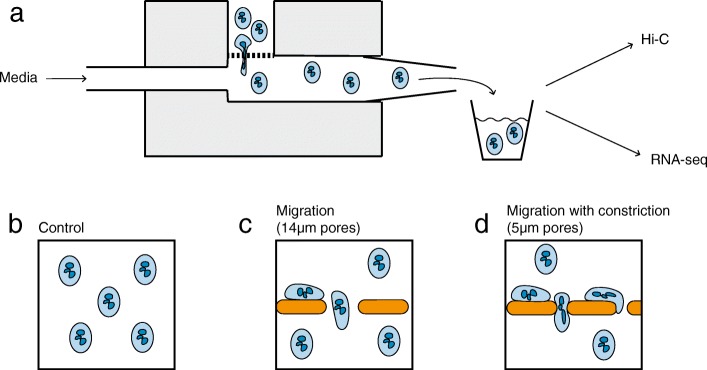
Background: Mammalian cells are flexible and can rapidly change shape when they contract, adhere, or migrate. The nucleus must be stiff enough to withstand cytoskeletal forces, but flexible enough to remodel as the cell changes shape. This is particularly important for cells migrating through confined spaces, where the nuclear shape must change in order to fit through a constriction. This occurs many times in the life cycle of a neutrophil, which must protect its chromatin from damage and disruption associated with migration. Here we characterized the effects of constricted migration in neutrophil-like cells.
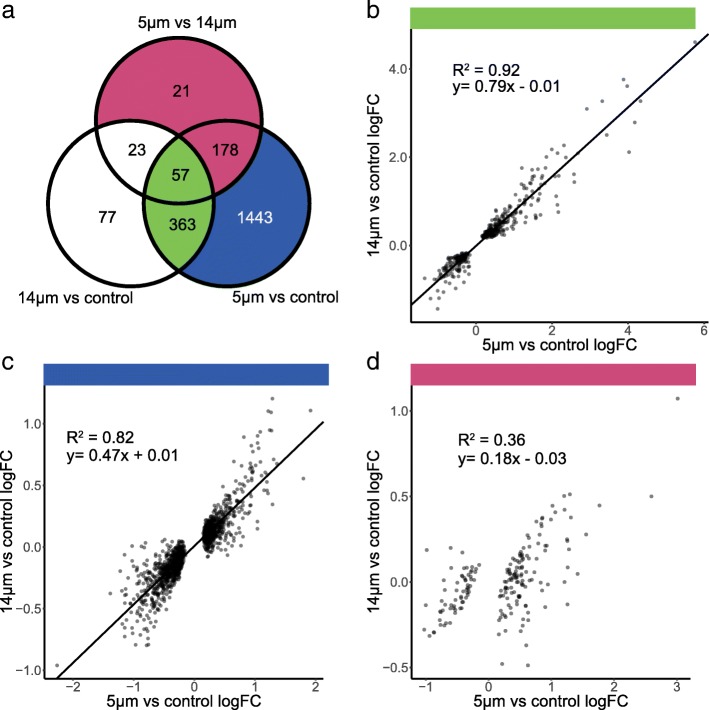
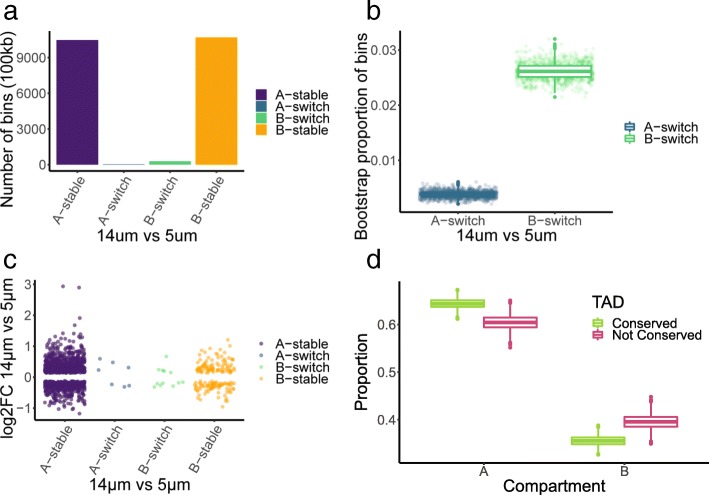
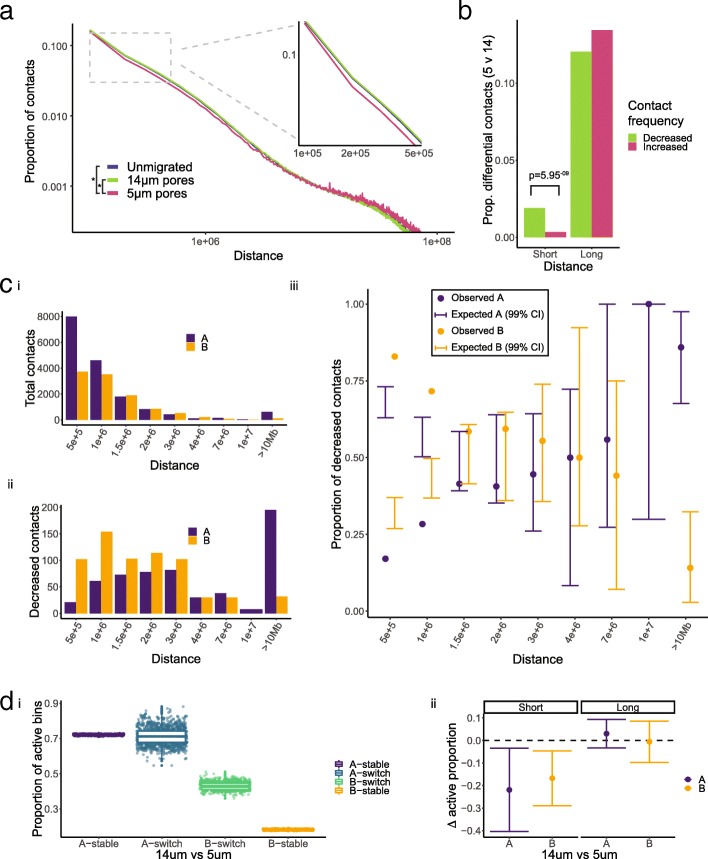
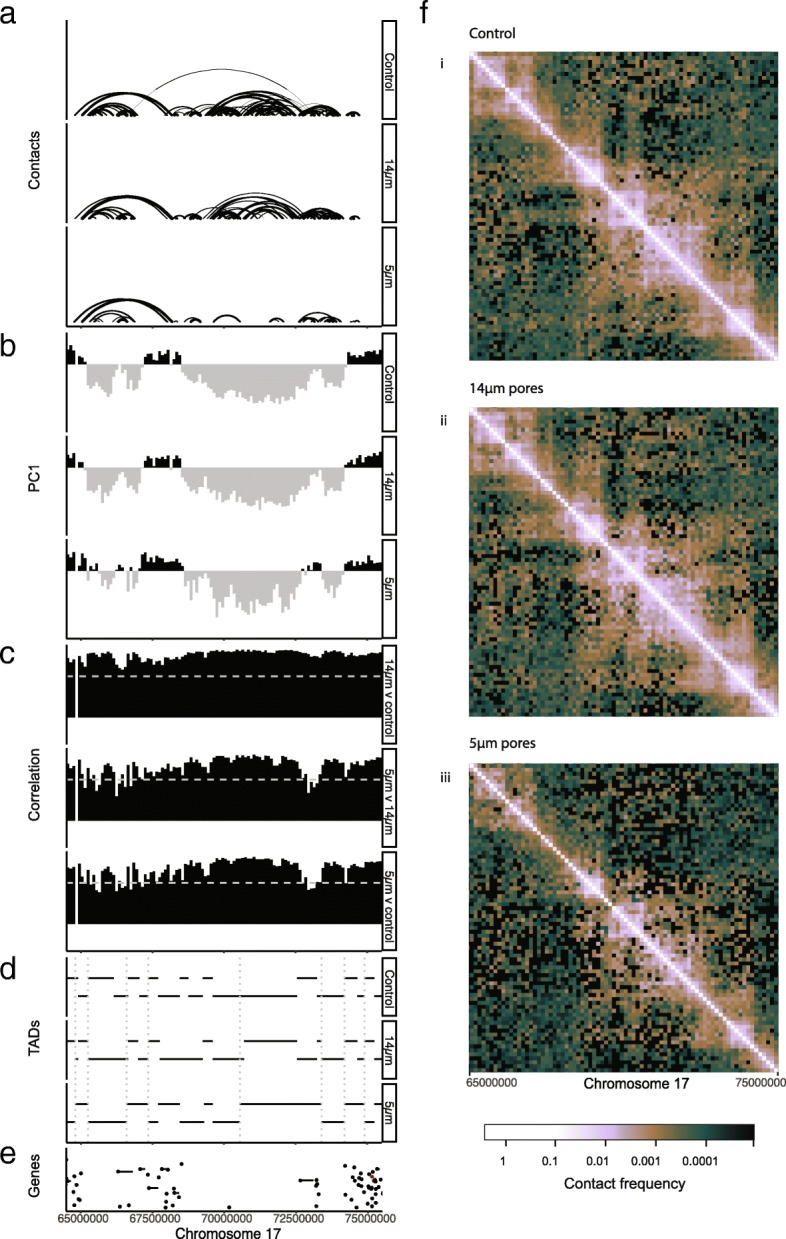
Results: Total RNA sequencing identified that migration of neutrophil-like cells through 5- or 14-μm pores was associated with changes in the transcript levels of inflammation and chemotaxis-related genes when compared to unmigrated cells. Differentially expressed transcripts specific to migration with constriction were enriched for groups of genes associated with cytoskeletal remodeling. Hi-C was used to capture the genome organization in control and migrated cells. Limited switching was observed between the active (A) and inactive (B) compartments after migration. However, global depletion of short-range contacts was observed following migration with constriction compared to migration without constriction. Regions with disrupted contacts, TADs, and compartments were enriched for inactive chromatin.
Conclusion: Short-range genome organization is preferentially altered in inactive chromatin, possibly protecting transcriptionally active contacts from the disruptive effects of migration with constriction. This is consistent with current hypotheses implicating heterochromatin as the mechanoresponsive form of chromatin. Further investigation concerning the contribution of heterochromatin to stiffness, flexibility, and protection of nuclear function will be important for understanding cell migration in relation to human health and disease.
Keywords: Chromatin conformation; Epigenetics; Heterochromatin; Hi-C; Immune; Mechanotransduction; Migration; Neutrophil; Nuclear remodeling; Transcription.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
Nuclear organization of active and inactive chromatin domains uncovered by chromosome conformation capture-on-chip (4C).Nat Genet. 2006 Nov;38(11):1348-54. doi: 10.1038/ng1896. Epub 2006 Oct 8. Nat Genet. 2006. PMID: 17033623
-
The Neutrophil Nucleus: An Important Influence on Neutrophil Migration and Function.Front Immunol. 2018 Dec 4;9:2867. doi: 10.3389/fimmu.2018.02867. eCollection 2018. Front Immunol. 2018. PMID: 30564248 Free PMC article. Review.
-
Confined migration induces heterochromatin formation and alters chromatin accessibility.iScience. 2022 Aug 17;25(9):104978. doi: 10.1016/j.isci.2022.104978. eCollection 2022 Sep 16. iScience. 2022. PMID: 36117991 Free PMC article.
-
Heterochromatin drives compartmentalization of inverted and conventional nuclei.Nature. 2019 Jun;570(7761):395-399. doi: 10.1038/s41586-019-1275-3. Epub 2019 Jun 5. Nature. 2019. PMID: 31168090 Free PMC article.
-
Emerging roles for the nucleus during neutrophil signal relay and NETosis.Curr Opin Cell Biol. 2020 Feb;62:135-143. doi: 10.1016/j.ceb.2019.10.007. Epub 2019 Dec 10. Curr Opin Cell Biol. 2020. PMID: 31835148 Review.
Cited by
-
Chromatin compaction during confined cell migration induces and reshapes nuclear condensates.Nat Commun. 2024 Nov 18;15(1):9964. doi: 10.1038/s41467-024-54120-5. Nat Commun. 2024. PMID: 39557835 Free PMC article.
-
Hyperosmotic stress: in situ chromatin phase separation.Nucleus. 2020 Dec;11(1):1-18. doi: 10.1080/19491034.2019.1710321. Nucleus. 2020. PMID: 31924112 Free PMC article.
-
Chromatin's physical properties shape the nucleus and its functions.Curr Opin Cell Biol. 2019 Jun;58:76-84. doi: 10.1016/j.ceb.2019.02.006. Epub 2019 Mar 16. Curr Opin Cell Biol. 2019. PMID: 30889417 Free PMC article. Review.
-
Constricted migration is associated with stable 3D genome structure differences in cancer cells.EMBO Rep. 2022 Oct 6;23(10):e52149. doi: 10.15252/embr.202052149. Epub 2022 Aug 15. EMBO Rep. 2022. PMID: 35969179 Free PMC article.
-
Lamina-associated domains: peripheral matters and internal affairs.Genome Biol. 2020 Apr 2;21(1):85. doi: 10.1186/s13059-020-02003-5. Genome Biol. 2020. PMID: 32241294 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
