

IKZF2 Drives Leukemia Stem Cell Self-Renewal and Inhibits Myeloid Differentiation
- PMID: 30472158
- PMCID: PMC6602096
- DOI: 10.1016/j.stem.2018.10.016
IKZF2 Drives Leukemia Stem Cell Self-Renewal and Inhibits Myeloid Differentiation
Abstract
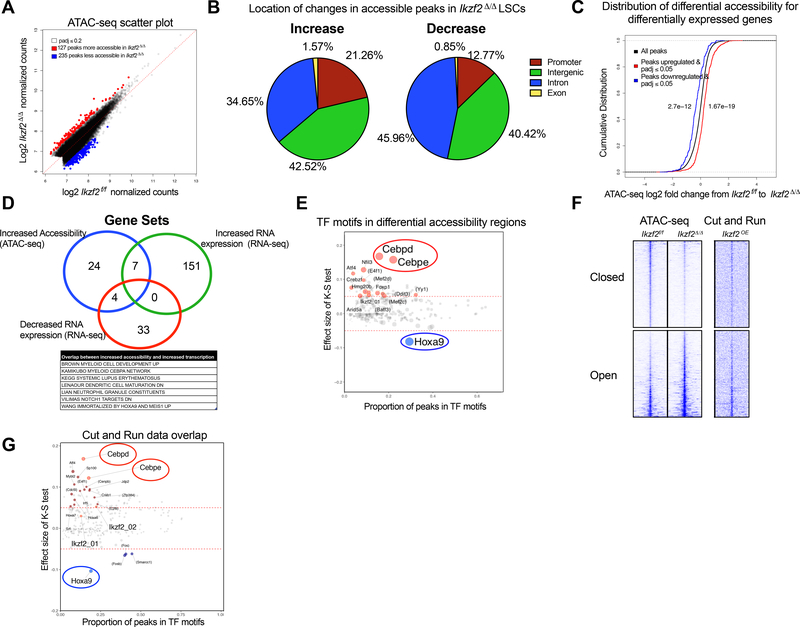
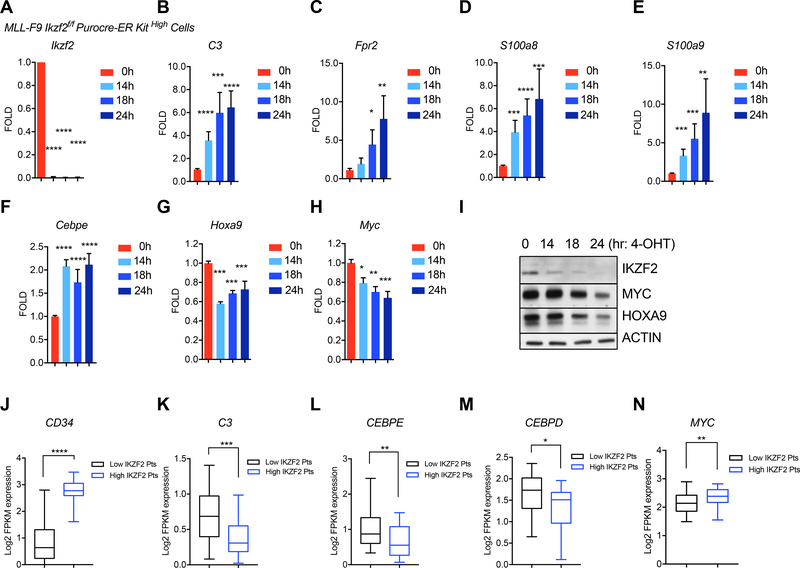
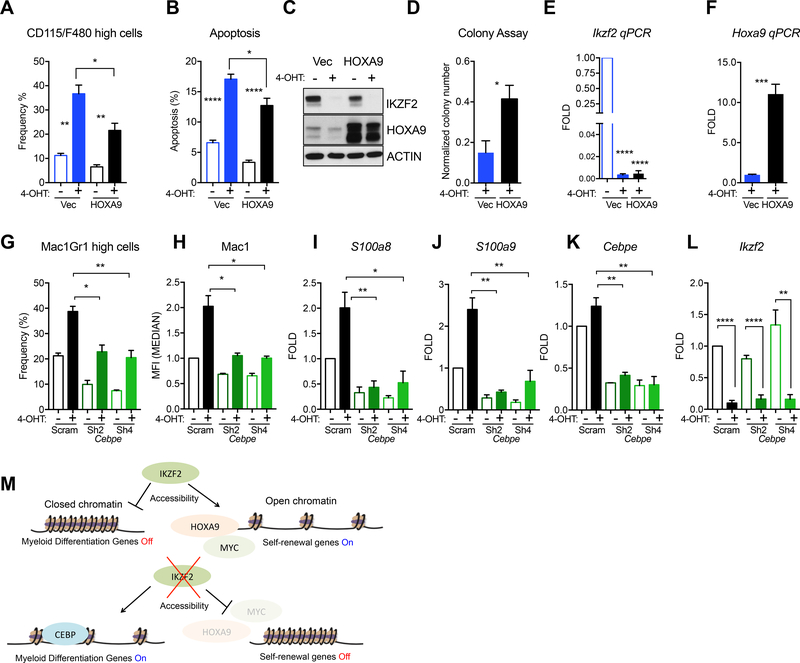
Leukemias exhibit a dysregulated developmental program mediated through both genetic and epigenetic mechanisms. Although IKZF2 is expressed in hematopoietic stem cells (HSCs), we found that it is dispensable for mouse and human HSC function. In contrast to its role as a tumor suppressor in hypodiploid B-acute lymphoblastic leukemia, we found that IKZF2 is required for myeloid leukemia. IKZF2 is highly expressed in leukemic stem cells (LSCs), and its deficiency results in defective LSC function. IKZF2 depletion in acute myeloid leukemia (AML) cells reduced colony formation, increased differentiation and apoptosis, and delayed leukemogenesis. Gene expression, chromatin accessibility, and direct IKZF2 binding in MLL-AF9 LSCs demonstrate that IKZF2 regulates a HOXA9 self-renewal gene expression program and inhibits a C/EBP-driven differentiation program. Ectopic HOXA9 expression and CEBPE depletion rescued the effects of IKZF2 depletion. Thus, our study shows that IKZF2 regulates the AML LSC program and provides a rationale to therapeutically target IKZF2 in myeloid leukemia.
Keywords: C/EBP; HOXA9; IKZF2; leukemic stem cells.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
S.A.A. consults for Epizyme Inc, Imago Biosciences, Cyteir Therapeutics, C4 Therapeutics, Syros Pharmaceuticals and Accent Therapeutics. S.A.A. receives research support from Janssen, Novartis, and AstraZeneca.
Figures
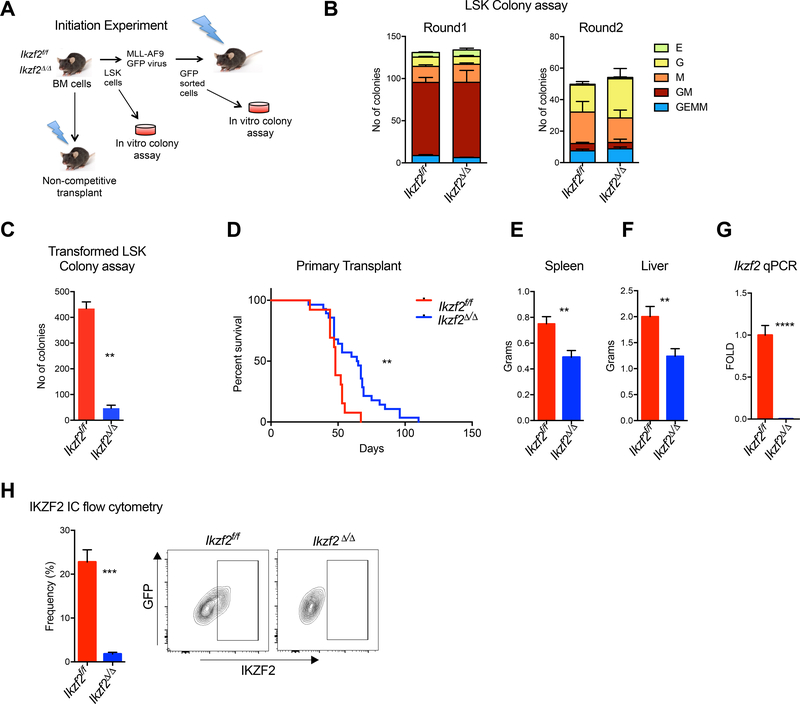
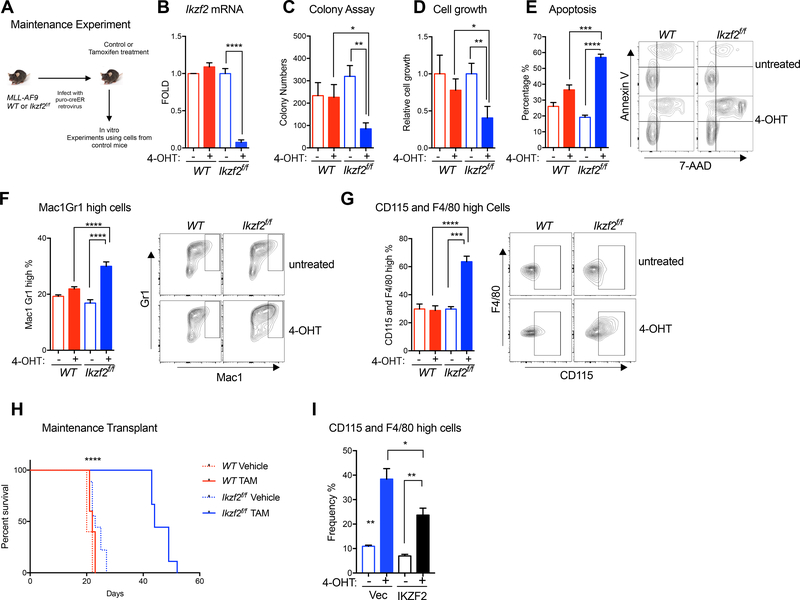
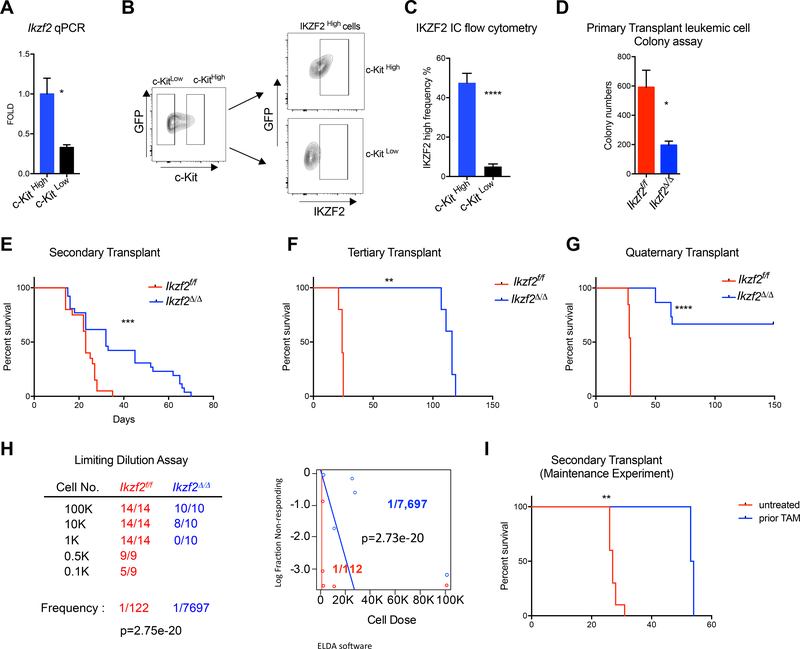
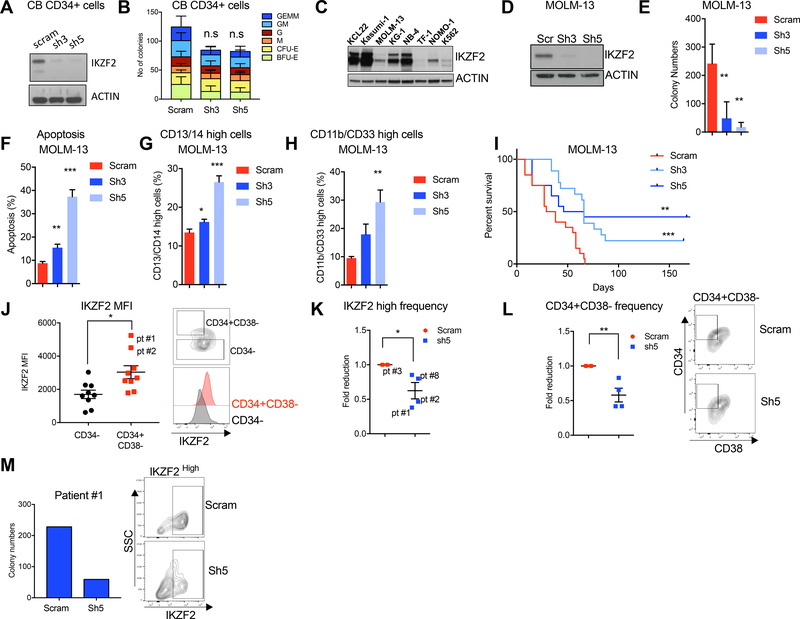
Comment in
-
The Making of a Leukemic Stem Cell: A Novel Role for IKZF2 in AML Stemness and Differentiation.Cell Stem Cell. 2019 Jan 3;24(1):5-6. doi: 10.1016/j.stem.2018.12.007. Cell Stem Cell. 2019. PMID: 30609399
Similar articles
-
Single-Cell Gene Expression Analyses Reveal Distinct Self-Renewing and Proliferating Subsets in the Leukemia Stem Cell Compartment in Acute Myeloid Leukemia.Cancer Res. 2020 Feb 1;80(3):458-470. doi: 10.1158/0008-5472.CAN-18-2932. Epub 2019 Nov 29. Cancer Res. 2020. PMID: 31784425 Free PMC article.
-
MLL-AF9- and HOXA9-mediated acute myeloid leukemia stem cell self-renewal requires JMJD1C.J Clin Invest. 2016 Mar 1;126(3):997-1011. doi: 10.1172/JCI82978. Epub 2016 Feb 15. J Clin Invest. 2016. PMID: 26878175 Free PMC article.
-
Musashi2 sustains the mixed-lineage leukemia-driven stem cell regulatory program.J Clin Invest. 2015 Mar 2;125(3):1286-98. doi: 10.1172/JCI78440. Epub 2015 Feb 9. J Clin Invest. 2015. PMID: 25664853 Free PMC article.
-
The cell of origin and the leukemia stem cell in acute myeloid leukemia.Genes Chromosomes Cancer. 2019 Dec;58(12):850-858. doi: 10.1002/gcc.22805. Epub 2019 Aug 31. Genes Chromosomes Cancer. 2019. PMID: 31471945 Review.
-
Understanding of leukemic stem cells and their clinical implications.Mol Cancer. 2017 Jan 30;16(1):2. doi: 10.1186/s12943-016-0574-7. Mol Cancer. 2017. PMID: 28137304 Free PMC article. Review.
Cited by
-
Design and Development of IKZF2 and CK1α Dual Degraders.J Med Chem. 2023 Dec 28;66(24):16953-16979. doi: 10.1021/acs.jmedchem.3c01736. Epub 2023 Dec 12. J Med Chem. 2023. PMID: 38085607 Free PMC article.
-
DNMT3A Harboring Leukemia-Associated Mutations Directs Sensitivity to DNA Damage at Replication Forks.Clin Cancer Res. 2022 Feb 15;28(4):756-769. doi: 10.1158/1078-0432.CCR-21-2863. Clin Cancer Res. 2022. PMID: 34716195 Free PMC article.
-
Identification of chromatin states during zebrafish gastrulation using CUT&RUN and CUT&Tag.Dev Dyn. 2022 Apr;251(4):729-742. doi: 10.1002/dvdy.430. Epub 2021 Oct 23. Dev Dyn. 2022. PMID: 34647658 Free PMC article.
-
miR-30e-5p regulates leukemia stem cell self-renewal through the Cyb561/ROS signaling pathway.Haematologica. 2024 Feb 1;109(2):411-421. doi: 10.3324/haematol.2023.282837. Haematologica. 2024. PMID: 37584287 Free PMC article.
-
Low doses of IFN-γ maintain self-renewal of leukemia stem cells in acute myeloid leukemia.Oncogene. 2023 Dec;42(50):3657-3669. doi: 10.1038/s41388-023-02874-5. Epub 2023 Oct 23. Oncogene. 2023. PMID: 37872214
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
