

Chemical biology probes of mammalian GLUT structure and function
- PMID: 30459202
- PMCID: PMC6243331
- DOI: 10.1042/BCJ20170677
Chemical biology probes of mammalian GLUT structure and function
Abstract
The structure and function of glucose transporters of the mammalian GLUT family of proteins has been studied over many decades, and the proteins have fascinated numerous research groups over this time. This interest is related to the importance of the GLUTs as archetypical membrane transport facilitators, as key limiters of the supply of glucose to cell metabolism, as targets of cell insulin and exercise signalling and of regulated membrane traffic, and as potential drug targets to combat cancer and metabolic diseases such as type 2 diabetes and obesity. This review focusses on the use of chemical biology approaches and sugar analogue probes to study these important proteins.
Keywords: GLUT proteins; GLUT4 translocation; insulin action.
© 2018 The Author(s).
Conflict of interest statement
The Author declares that there are no competing interests associated with this manuscript.
Figures
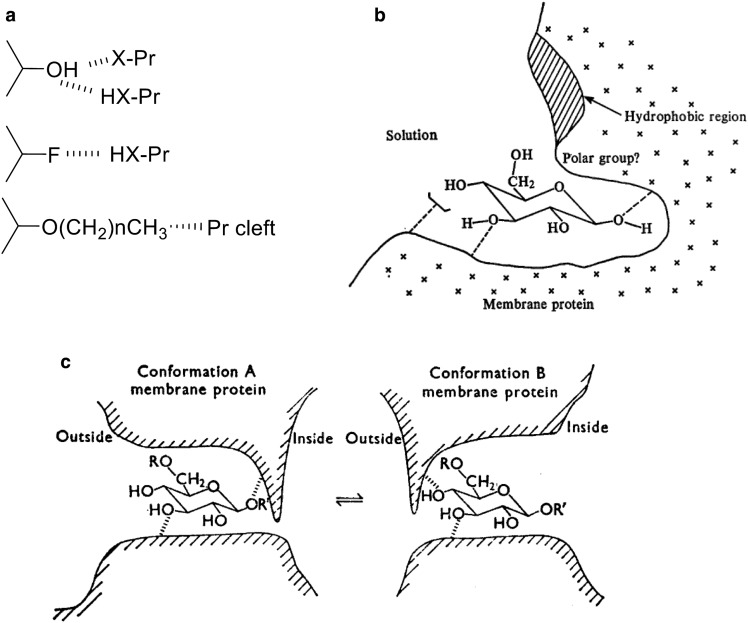
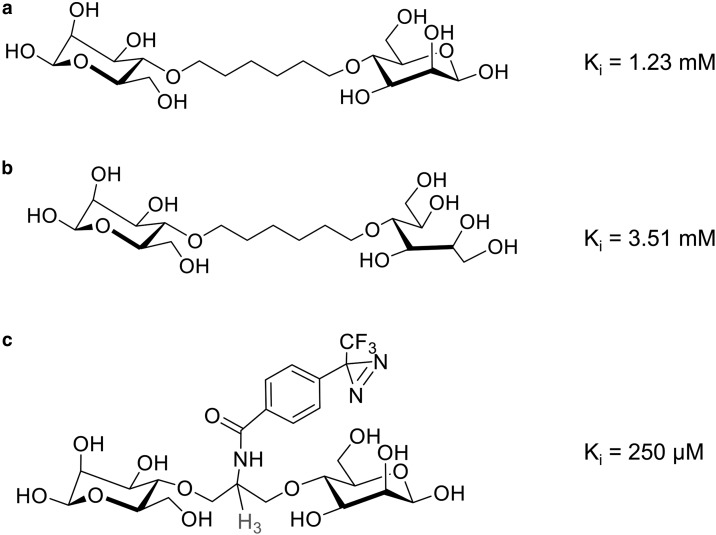
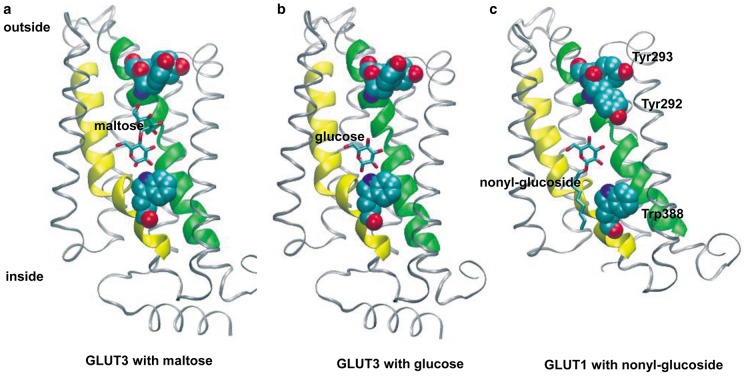
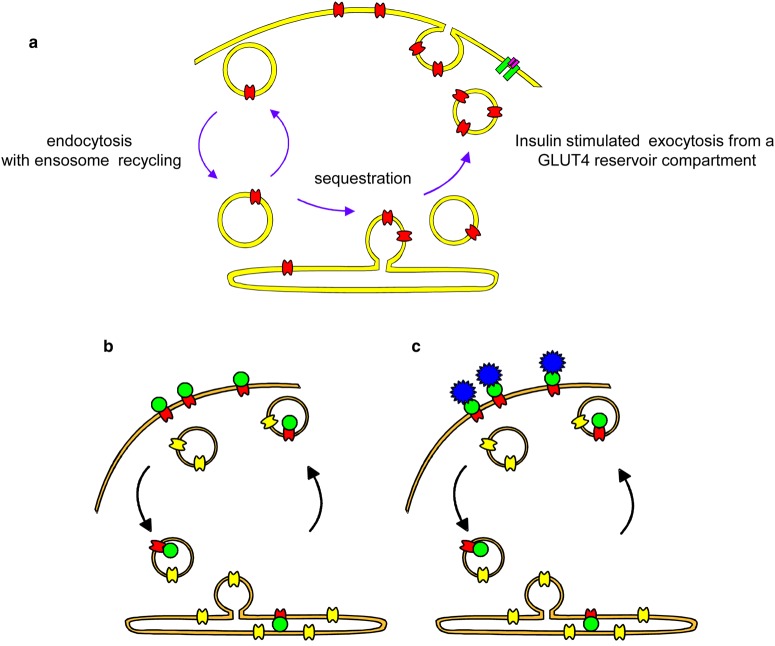
Similar articles
-
Structure, function and regulation of mammalian glucose transporters of the SLC2 family.Pflugers Arch. 2020 Sep;472(9):1155-1175. doi: 10.1007/s00424-020-02411-3. Epub 2020 Jun 26. Pflugers Arch. 2020. PMID: 32591905 Free PMC article. Review.
-
Molecular Tools for Facilitative Carbohydrate Transporters (Gluts).Chembiochem. 2017 Sep 19;18(18):1774-1788. doi: 10.1002/cbic.201700221. Epub 2017 Aug 1. Chembiochem. 2017. PMID: 28636761 Review.
-
Importance of GLUT Transporters in Disease Diagnosis and Treatment.Int J Mol Sci. 2022 Aug 4;23(15):8698. doi: 10.3390/ijms23158698. Int J Mol Sci. 2022. PMID: 35955833 Free PMC article. Review.
-
A Glimpse of Membrane Transport through Structures-Advances in the Structural Biology of the GLUT Glucose Transporters.J Mol Biol. 2017 Aug 18;429(17):2710-2725. doi: 10.1016/j.jmb.2017.07.009. Epub 2017 Jul 26. J Mol Biol. 2017. PMID: 28756087 Review.
-
Diabetes Alters the Expression and Translocation of the Insulin-Sensitive Glucose Transporters 4 and 8 in the Atria.PLoS One. 2015 Dec 31;10(12):e0146033. doi: 10.1371/journal.pone.0146033. eCollection 2015. PLoS One. 2015. PMID: 26720696 Free PMC article.
Cited by
-
Visualization of GLUT1 Trafficking in Live Cancer Cells by the Use of a Dual-Fluorescence Reporter.ACS Omega. 2020 Jun 23;5(26):15911-15921. doi: 10.1021/acsomega.0c01054. eCollection 2020 Jul 7. ACS Omega. 2020. PMID: 32656411 Free PMC article.
-
Structures and General Transport Mechanisms by the Major Facilitator Superfamily (MFS).Chem Rev. 2021 May 12;121(9):5289-5335. doi: 10.1021/acs.chemrev.0c00983. Epub 2021 Apr 22. Chem Rev. 2021. PMID: 33886296 Free PMC article.
-
Apigenin Suppresses the Warburg Effect and Stem-like Properties in SOSP-9607 Cells by Inactivating the PI3K/Akt/mTOR Signaling Pathway.Evid Based Complement Alternat Med. 2022 Mar 9;2022:3983637. doi: 10.1155/2022/3983637. eCollection 2022. Evid Based Complement Alternat Med. 2022. PMID: 35310040 Free PMC article.
-
The Arabidopsis AtSWEET13 transporter discriminates sugars by selective facial and positional substrate recognition.Commun Biol. 2024 Jun 24;7(1):764. doi: 10.1038/s42003-024-06291-6. Commun Biol. 2024. PMID: 38914639 Free PMC article.
-
Expression and Prognostic Value of Glucose Transporter 3 in Diffuse Large B Cell Lymphoma.Onco Targets Ther. 2022 Feb 25;15:181-191. doi: 10.2147/OTT.S338826. eCollection 2022. Onco Targets Ther. 2022. PMID: 35250277 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical