

Elicitation of Protective Antibodies against 20 Years of Future H3N2 Cocirculating Influenza Virus Variants in Ferrets Preimmune to Historical H3N2 Influenza Viruses
- PMID: 30429350
- PMCID: PMC6340030
- DOI: 10.1128/JVI.00946-18
Elicitation of Protective Antibodies against 20 Years of Future H3N2 Cocirculating Influenza Virus Variants in Ferrets Preimmune to Historical H3N2 Influenza Viruses
Abstract
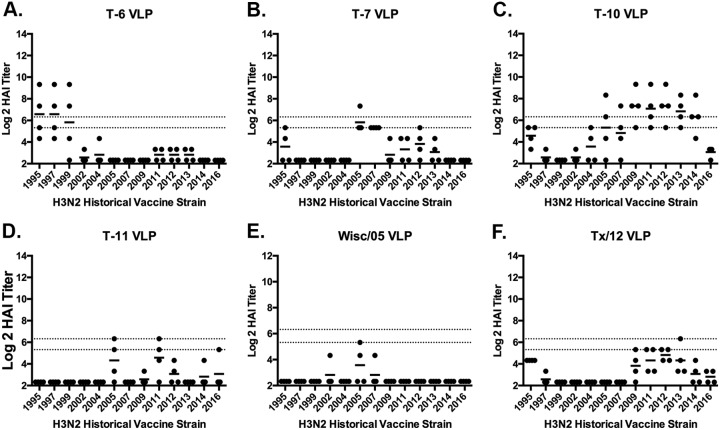
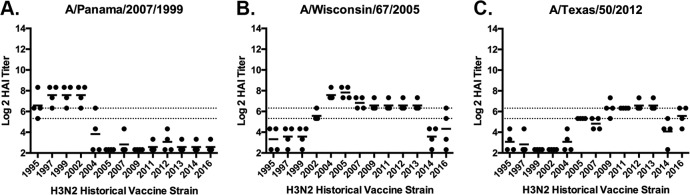
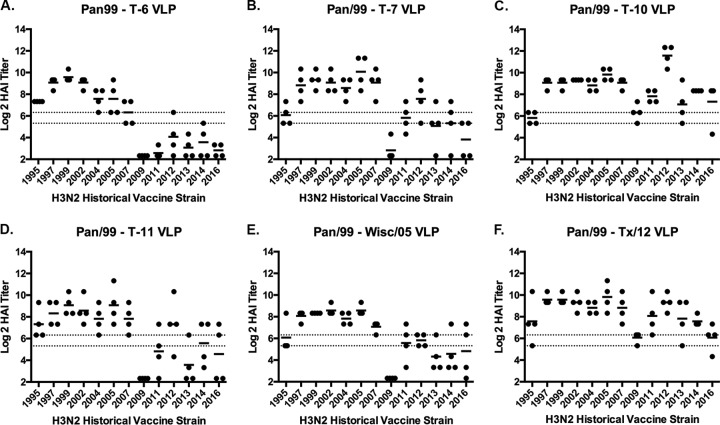
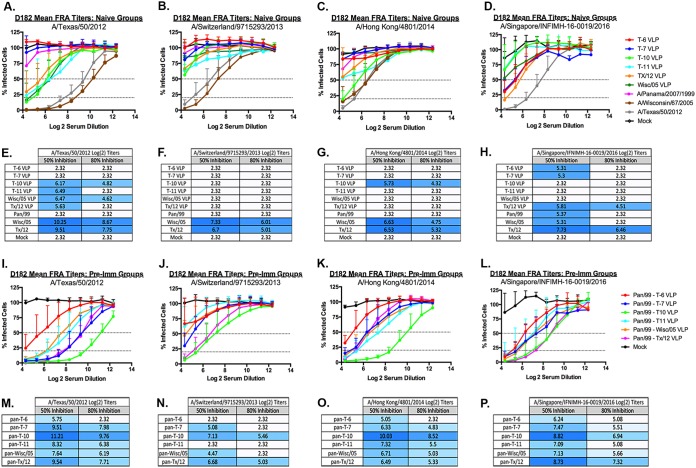
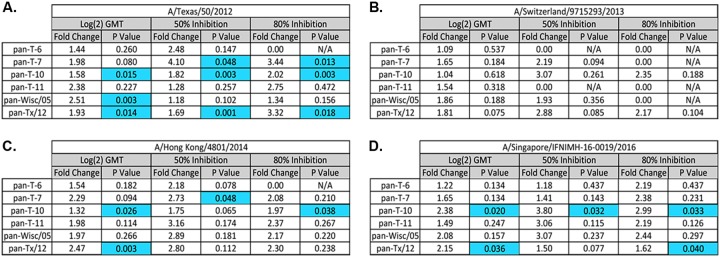
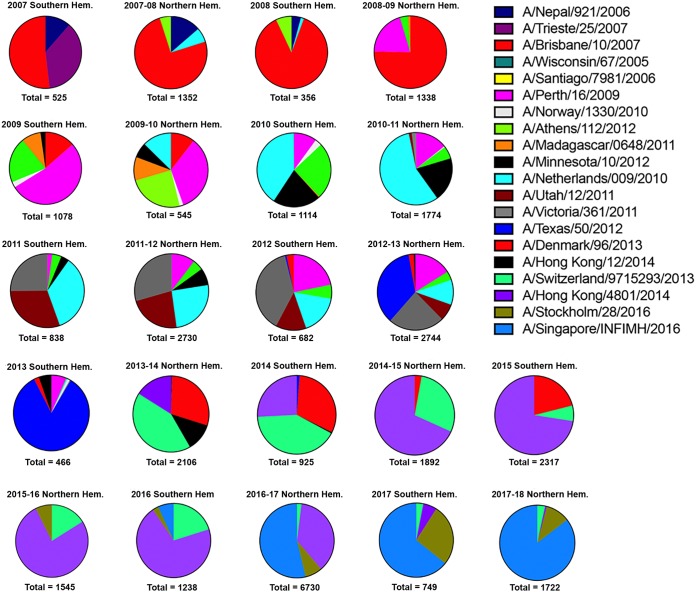
The vast majority of people already have preexisting immune responses to influenza viruses from one or more subtypes. However, almost all preclinical studies evaluate new influenza vaccine candidates in immunologically naive animals. Recently, our group demonstrated that priming naive ferrets with broadly reactive H1 COBRA HA-based vaccines boosted preexisting antibodies induced by wild-type H1N1 virus infections. These H1 COBRA hemagglutinin (HA) antigens induced antibodies with HAI activity against multiple antigenically different H1N1 viral variants. In this study, ferrets, preimmune to historical H3N2 viruses, were vaccinated with virus-like particle (VLP) vaccines expressing either an HA from a wild-type H3 influenza virus or a COBRA H3 HA antigen (T6, T7, T10, or T11). The elicited antisera had the ability to neutralize virus infection against either a panel of viruses representing vaccine strains selected by the World Health Organization or a set of viral variants that cocirculated during the same time period. Preimmune animals vaccinated with H3 COBRA T10 HA antigen elicited sera with higher hemagglutination inhibition (HAI) antibody titers than antisera elicited by VLP vaccines with wild-type HA VLPs in preimmune ferrets. However, while the T11 COBRA vaccine did not elicit HAI activity, the elicited antibodies did neutralize antigenically distinct H3N2 influenza viruses. Overall, H3 COBRA-based HA vaccines were able to neutralize both historical H3 and contemporary, as well as future, H3N2 viruses with higher titers than vaccines with wild-type H3 HA antigens. This is the first report demonstrating the effectiveness of a broadly reactive H3N3 vaccine in a preimmune ferret model.IMPORTANCE After exposure to influenza virus, the host generates neutralizing anti-hemagglutinin (anti-HA) antibodies against that specific infecting influenza strain. These antibodies can also neutralize some, but not all, cocirculating strains. The goal of next-generation influenza vaccines, such as HA head-based COBRA, is to stimulate broadly protective neutralizing antibodies against all strains circulating within a subtype, in particular those that persist over multiple influenza seasons, without requiring an update to the vaccine. To mimic the human condition, COBRA HA virus-like particle vaccines were tested in ferrets that were previously exposed to historical H3N2 influenza viruses. In this model, these vaccines elicited broadly protective antibodies that neutralized cocirculating H3N2 influenza viruses isolated over a 20-year period. This is the first study to show the effectiveness of H3N3 COBRA HA vaccines in a host with preexisting immunity to influenza.
Keywords: COBRA; H3N2; broadly protective vaccine; ferrets; hemagglutination inhibition; influenza; neutralization.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Elicitation of Protective Antibodies against a Broad Panel of H1N1 Viruses in Ferrets Preimmune to Historical H1N1 Influenza Viruses.J Virol. 2017 Nov 30;91(24):e01283-17. doi: 10.1128/JVI.01283-17. Print 2017 Dec 15. J Virol. 2017. Retraction in: J Virol. 2024 Jul 23;98(7):e0183223. doi: 10.1128/jvi.01832-23 PMID: 28978709 Free PMC article. Retracted.
-
Computationally Optimized Broadly Reactive Hemagglutinin Elicits Hemagglutination Inhibition Antibodies against a Panel of H3N2 Influenza Virus Cocirculating Variants.J Virol. 2017 Nov 30;91(24):e01581-17. doi: 10.1128/JVI.01581-17. Print 2017 Dec 15. J Virol. 2017. PMID: 28978710 Free PMC article.
-
Broadly Reactive H2 Hemagglutinin Vaccines Elicit Cross-Reactive Antibodies in Ferrets Preimmune to Seasonal Influenza A Viruses.mSphere. 2021 Mar 10;6(2):e00052-21. doi: 10.1128/mSphere.00052-21. mSphere. 2021. PMID: 33692193 Free PMC article.
-
Next-Generation Influenza HA Immunogens and Adjuvants in Pursuit of a Broadly Protective Vaccine.Viruses. 2021 Mar 24;13(4):546. doi: 10.3390/v13040546. Viruses. 2021. PMID: 33805245 Free PMC article. Review.
-
Influenza Hemagglutinin Structures and Antibody Recognition.Cold Spring Harb Perspect Med. 2020 Aug 3;10(8):a038778. doi: 10.1101/cshperspect.a038778. Cold Spring Harb Perspect Med. 2020. PMID: 31871236 Free PMC article. Review.
Cited by
-
Strategies Targeting Hemagglutinin as a Universal Influenza Vaccine.Vaccines (Basel). 2021 Mar 13;9(3):257. doi: 10.3390/vaccines9030257. Vaccines (Basel). 2021. PMID: 33805749 Free PMC article. Review.
-
Interaction Between Virus-Like Particles (VLPs) and Pattern Recognition Receptors (PRRs) From Dendritic Cells (DCs): Toward Better Engineering of VLPs.Front Immunol. 2020 Jun 9;11:1100. doi: 10.3389/fimmu.2020.01100. eCollection 2020. Front Immunol. 2020. PMID: 32582186 Free PMC article. Review.
-
Differential Recognition of Computationally Optimized H3 Hemagglutinin Influenza Vaccine Candidates by Human Antibodies.J Virol. 2022 Aug 24;96(16):e0089622. doi: 10.1128/jvi.00896-22. Epub 2022 Aug 2. J Virol. 2022. PMID: 35916534 Free PMC article.
-
Immune Response in Mice Immunized with Chimeric H1 Antigens.Vaccines (Basel). 2021 Oct 15;9(10):1182. doi: 10.3390/vaccines9101182. Vaccines (Basel). 2021. PMID: 34696290 Free PMC article.
-
Pre-existing heterosubtypic immunity provides a barrier to airborne transmission of influenza viruses.PLoS Pathog. 2021 Feb 18;17(2):e1009273. doi: 10.1371/journal.ppat.1009273. eCollection 2021 Feb. PLoS Pathog. 2021. PMID: 33600489 Free PMC article.
References
-
- Flannery B, Chung JR, Belongia EA, McLean HQ, Gaglani M, Murthy K, Zimmerman RK, Nowalk MP, Jackson ML, Jackson LA, Monto AS, Martin ET, Foust A, Sessions W, Berman S, Barnes JR, Spencer S, Fry AM. 2018. Interim estimates of 2017–18 seasonal influenza vaccine effectiveness—United States, February 2018. MMWR Morb Mortal Wkly Rep 67:180–185. doi:10.15585/mmwr.mm6706a2. - DOI - PMC - PubMed
-
- Skowronski DM, Chambers C, De Serres G, Dickinson JA, Winter AL, Hickman R, Chan T, Jassem AN, Drews SJ, Charest H, Gubbay JB, Bastien N, Li Y, Krajden M. 2018. Early season cocirculation of influenza A(H3N2) and B(Yamagata): interim estimates of 2017/18 vaccine effectiveness, Canada. Euro Surveill 23. doi:10.2807/1560-7917.ES.2018.23.5.18-00035. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
