

Mutations in the Spike Protein of Middle East Respiratory Syndrome Coronavirus Transmitted in Korea Increase Resistance to Antibody-Mediated Neutralization
- PMID: 30404801
- PMCID: PMC6321919
- DOI: 10.1128/JVI.01381-18
Mutations in the Spike Protein of Middle East Respiratory Syndrome Coronavirus Transmitted in Korea Increase Resistance to Antibody-Mediated Neutralization
Abstract
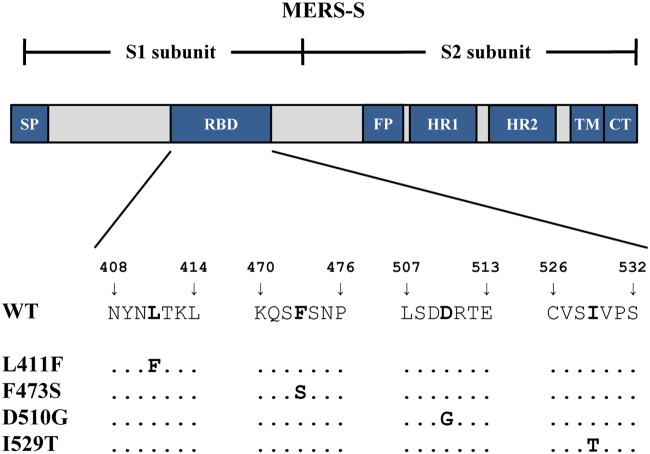
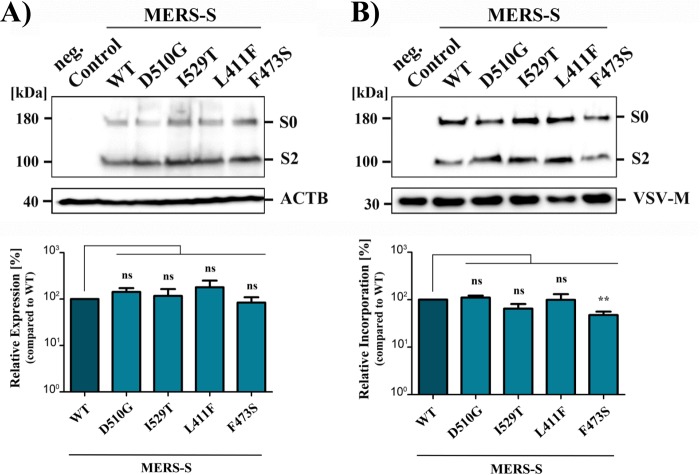
Middle East respiratory syndrome coronavirus (MERS-CoV) poses a threat to public health. The virus is endemic in the Middle East but can be transmitted to other countries by travel activity. The introduction of MERS-CoV into the Republic of Korea by an infected traveler resulted in a hospital outbreak of MERS that entailed 186 cases and 38 deaths. The MERS-CoV spike (S) protein binds to the cellular protein DPP4 via its receptor binding domain (RBD) and mediates viral entry into target cells. During the MERS outbreak in Korea, emergence and spread of viral variants that harbored mutations in the RBD, D510G and I529T, was observed. Counterintuitively, these mutations were found to reduce DPP4 binding and viral entry into target cells. In this study, we investigated whether they also exerted proviral effects. We confirm that changes D510G and I529T reduce S protein binding to DPP4 but show that this reduction only translates into diminished viral entry when expression of DPP4 on target cells is low. Neither mutation modulated S protein binding to sialic acids, S protein activation by host cell proteases, or inhibition of S protein-driven entry by interferon-induced transmembrane proteins. In contrast, changes D510G and I529T increased resistance of S protein-driven entry to neutralization by monoclonal antibodies and sera from MERS patients. These findings indicate that MERS-CoV variants with reduced neutralization sensitivity were transmitted during the Korean outbreak and that the responsible mutations were compatible with robust infection of cells expressing high levels of DPP4.IMPORTANCE MERS-CoV has pandemic potential, and it is important to identify mutations in viral proteins that might augment viral spread. In the course of a large hospital outbreak of MERS in the Republic of Korea in 2015, the spread of a viral variant that contained mutations in the viral spike protein was observed. These mutations were found to reduce receptor binding and viral infectivity. However, it remained unclear whether they also exerted proviral effects. We demonstrate that these mutations reduce sensitivity to antibody-mediated neutralization and are compatible with robust infection of target cells expressing large amounts of the viral receptor DPP4.
Keywords: MERS; antibody; neutralization; spike; virus entry.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Spread of Mutant Middle East Respiratory Syndrome Coronavirus with Reduced Affinity to Human CD26 during the South Korean Outbreak.mBio. 2016 Mar 1;7(2):e00019. doi: 10.1128/mBio.00019-16. mBio. 2016. PMID: 26933050 Free PMC article.
-
Recombinant Receptor-Binding Domains of Multiple Middle East Respiratory Syndrome Coronaviruses (MERS-CoVs) Induce Cross-Neutralizing Antibodies against Divergent Human and Camel MERS-CoVs and Antibody Escape Mutants.J Virol. 2016 Dec 16;91(1):e01651-16. doi: 10.1128/JVI.01651-16. Print 2017 Jan 1. J Virol. 2016. PMID: 27795425 Free PMC article.
-
Identification of sialic acid-binding function for the Middle East respiratory syndrome coronavirus spike glycoprotein.Proc Natl Acad Sci U S A. 2017 Oct 3;114(40):E8508-E8517. doi: 10.1073/pnas.1712592114. Epub 2017 Sep 18. Proc Natl Acad Sci U S A. 2017. PMID: 28923942 Free PMC article.
-
MERS-CoV spike protein: a key target for antivirals.Expert Opin Ther Targets. 2017 Feb;21(2):131-143. doi: 10.1080/14728222.2017.1271415. Epub 2016 Dec 21. Expert Opin Ther Targets. 2017. PMID: 27936982 Free PMC article. Review.
-
[Development of peptidic MERS-CoV entry inhibitors].Yao Xue Xue Bao. 2015 Dec;50(12):1513-9. Yao Xue Xue Bao. 2015. PMID: 27169270 Review. Chinese.
Cited by
-
Evaluation of the Neutralizing Antibody STE90-C11 against SARS-CoV-2 Delta Infection and Its Recognition of Other Variants of Concerns.Viruses. 2023 Oct 25;15(11):2153. doi: 10.3390/v15112153. Viruses. 2023. PMID: 38005829 Free PMC article.
-
Targeting the DEAD-Box RNA Helicase eIF4A with Rocaglates-A Pan-Antiviral Strategy for Minimizing the Impact of Future RNA Virus Pandemics.Microorganisms. 2021 Mar 5;9(3):540. doi: 10.3390/microorganisms9030540. Microorganisms. 2021. PMID: 33807988 Free PMC article. Review.
-
Monitoring and immunogenicity of SARS-CoV-2 vaccination of laboratory rhesus monkeys (Macaca mulatta).Sci Rep. 2023 Feb 25;13(1):3274. doi: 10.1038/s41598-023-30473-7. Sci Rep. 2023. PMID: 36841887 Free PMC article.
-
Combining a Fusion Inhibitory Peptide Targeting the MERS-CoV S2 Protein HR1 Domain and a Neutralizing Antibody Specific for the S1 Protein Receptor-Binding Domain (RBD) Showed Potent Synergism against Pseudotyped MERS-CoV with or without Mutations in RBD.Viruses. 2019 Jan 6;11(1):31. doi: 10.3390/v11010031. Viruses. 2019. PMID: 30621343 Free PMC article.
-
Sequential Emergence and Wide Spread of Neutralization Escape Middle East Respiratory Syndrome Coronavirus Mutants, South Korea, 2015.Emerg Infect Dis. 2019 Jun;25(6):1161-1168. doi: 10.3201/eid2506.181722. Epub 2019 Jun 17. Emerg Infect Dis. 2019. PMID: 30900977 Free PMC article.
References
-
- Lai MMC, Perlman S, Anderson LJ. 2007. Coronaviridae, p 1305–1336. In Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 5th ed, vol 1 Lippincott, Williams & Wilkins, Philadelphia, PA.
-
- Haagmans BL, Al Dhahiry SH, Reusken CB, Raj VS, Galiano M, Myers R, Godeke GJ, Jonges M, Farag E, Diab A, Ghobashy H, Alhajri F, Al-Thani M, Al-Marri SA, Al Romaihi HE, Al Khal A, Bermingham A, Osterhaus AD, AlHajri MM, Koopmans MP. 2014. Middle East respiratory syndrome coronavirus in dromedary camels: an outbreak investigation. Lancet Infect Dis 14:140–145. doi:10.1016/S1473-3099(13)70690-X. - DOI - PMC - PubMed
-
- Reusken CBEM, Haagmans BL, Müller MA, Gutierrez C, Godeke G-J, Meyer B, Muth D, Raj VS, Smits-De Vries L, Corman VM, Drexler J-F, Smits SL, El Tahir YE, De Sousa R, van Beek J, Nowotny N, van Maanen K, Hidalgo-Hermoso E, Bosch B-J, Rottier P, Osterhaus A, Gortázar-Schmidt C, Drosten C, Koopmans MPG. 2013. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: a comparative serological study. Lancet Infect Dis 13:859–866. doi:10.1016/S1473-3099(13)70164-6. - DOI - PMC - PubMed
-
- World Health Organization. 2018. Middle East respiratory syndrome coronavirus (MERS-CoV)—Saudi Arabia. http://www.emro.who.int/pandemic-epidemic-diseases/mers-cov/mers-situati.... Accessed 27 July 2018.
-
- Assiri A, McGeer A, Perl TM, Price CS, Al Rabeeah AA, Cummings DA, Alabdullatif ZN, Assad M, Almulhim A, Makhdoom H, Madani H, Alhakeem R, Al-Tawfiq JA, Cotten M, Watson SJ, Kellam P, Zumla AI, Memish ZA, KSA MERS-CoV Investigation Team . 2013. Hospital outbreak of Middle East respiratory syndrome coronavirus. N Engl J Med 369:407–416. doi:10.1056/NEJMoa1306742. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
