

Role of oligouridylation in normal metabolism and regulated degradation of mammalian histone mRNAs
- PMID: 30397106
- PMCID: PMC6232589
- DOI: 10.1098/rstb.2018.0170
Role of oligouridylation in normal metabolism and regulated degradation of mammalian histone mRNAs
Abstract
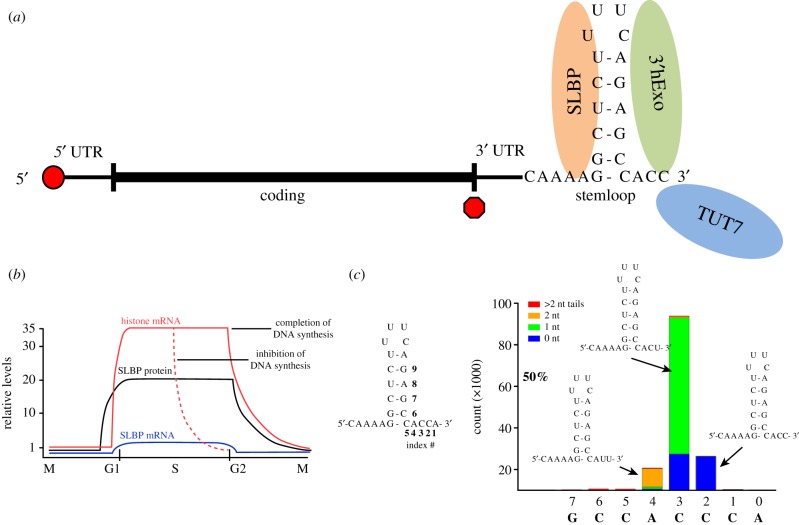
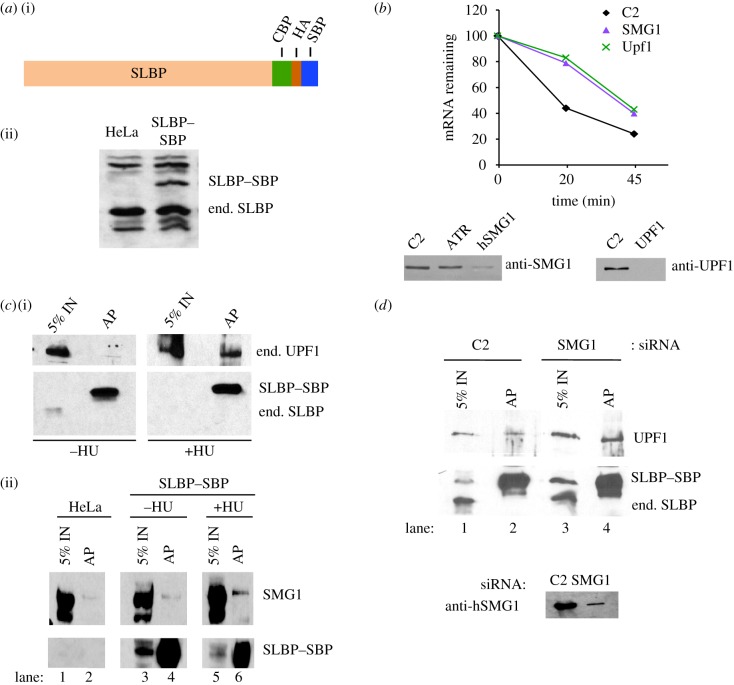
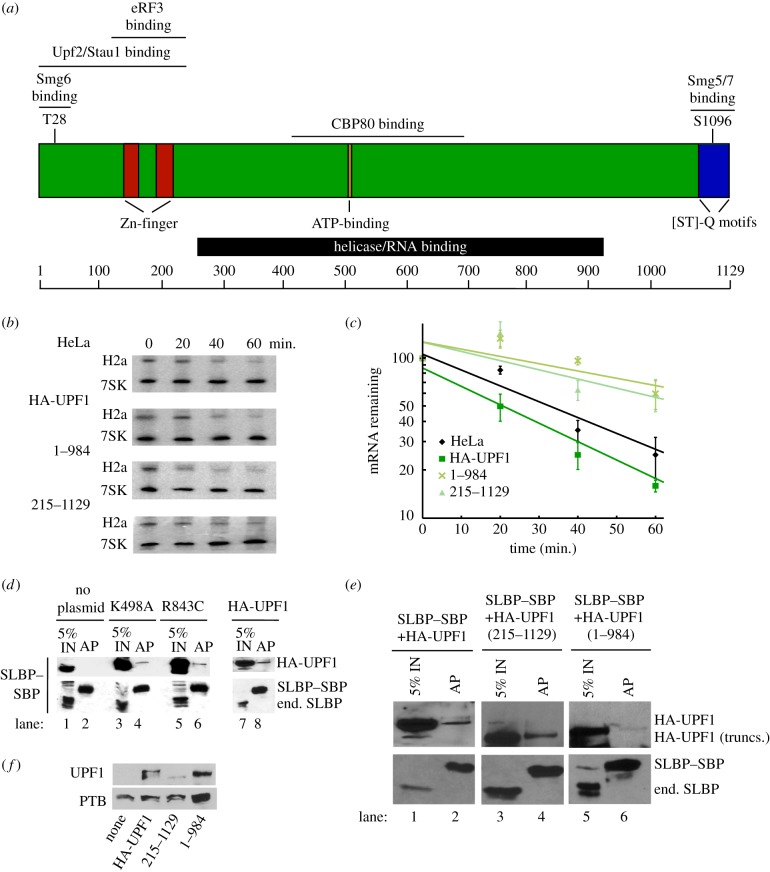
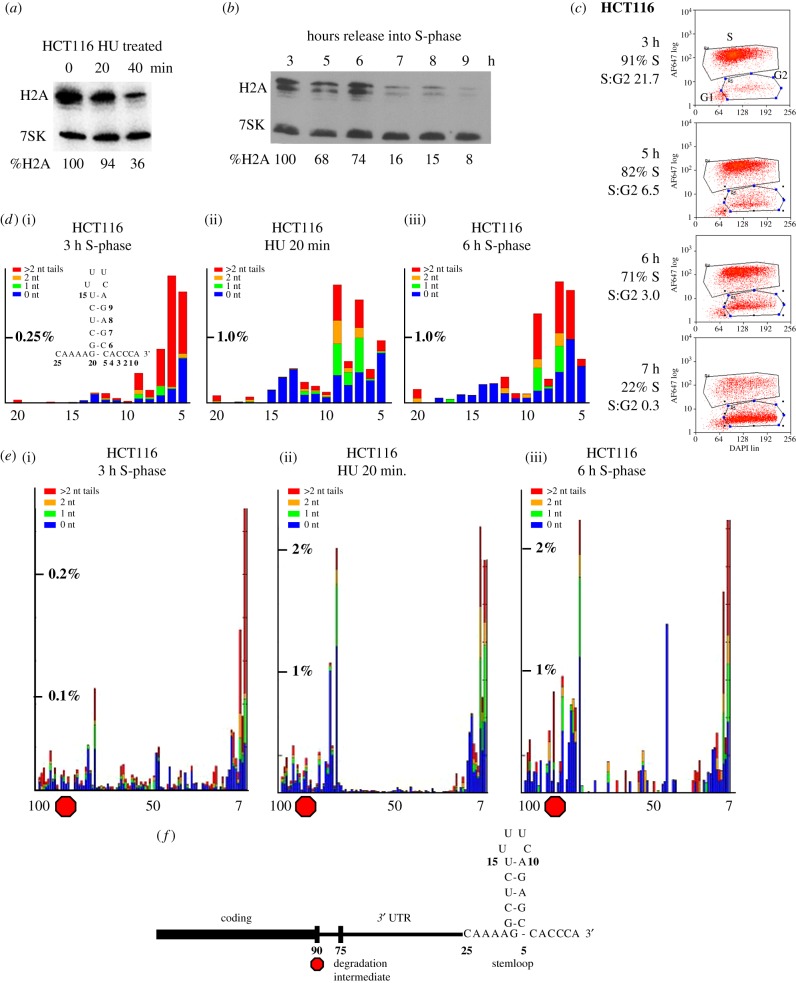
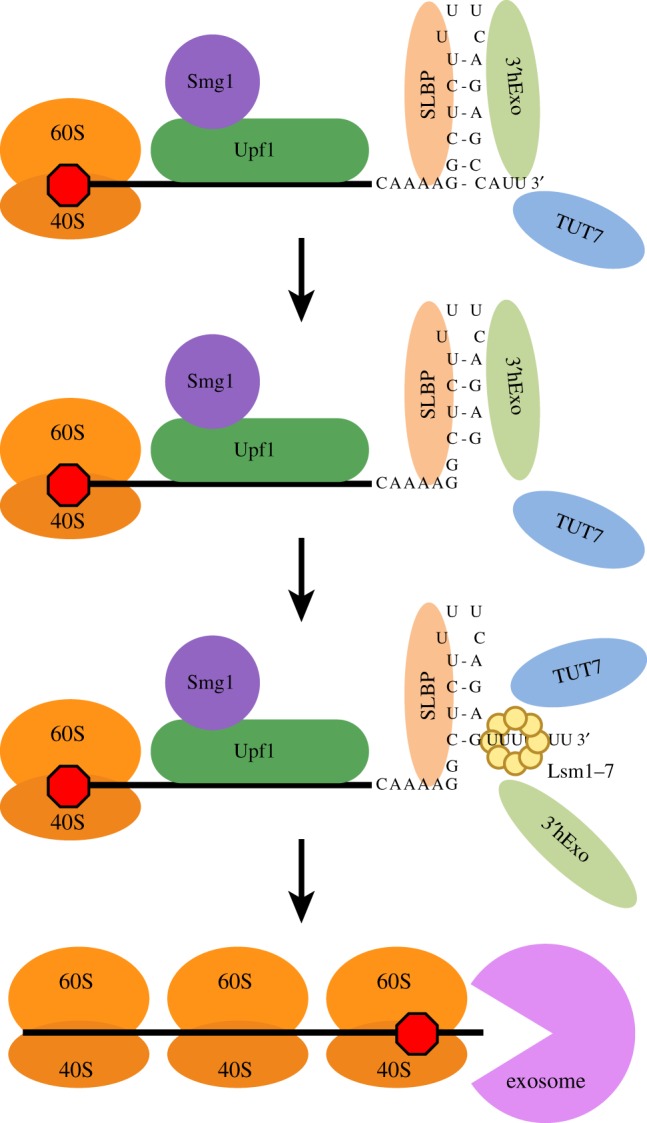
Metazoan replication-dependent histone mRNAs are the only known cellular mRNAs that are not polyadenylated. Histone mRNAs are present in large amounts only in S-phase cells, and their levels are coordinately regulated with the rate of DNA replication. In mammals, the stemloop at the 3' end of histone mRNA is bound to stemloop binding protein, a protein required for both synthesis and degradation of histone mRNA, and an exonuclease, 3'hExo (ERI1). Histone mRNAs are rapidly degraded when DNA synthesis is inhibited in S-phase cells and at the end of S-phase. Upf1 is also required for rapid degradation of histone mRNA as is the S-phase checkpoint. We report that Smg1 is required for histone mRNA degradation when DNA replication is inhibited, suggesting it is the PI-like kinase that activates Upf1 for histone mRNA degradation. We also show that some mutant Upf1 proteins are recruited to histone mRNAs when DNA replication is inhibited and act as dominant negative factors in histone mRNA degradation. We report that the pathway of rapid histone mRNA degradation when DNA replication is inhibited in S-phase cells that are activating the S-phase checkpoint is similar to the pathway of rapid degradation of histone mRNA at the end of S-phase.This article is part of the theme issue '5' and 3' modifications controlling RNA degradation'.
Keywords: RNA degradation; cell cycle; histone mRNA; uridylation.
© 2018 The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
TUT7 catalyzes the uridylation of the 3' end for rapid degradation of histone mRNA.RNA. 2016 Nov;22(11):1673-1688. doi: 10.1261/rna.058107.116. Epub 2016 Sep 8. RNA. 2016. PMID: 27609902 Free PMC article.
-
Knockouts of TUT7 and 3'hExo show that they cooperate in histone mRNA maintenance and degradation.RNA. 2022 Nov;28(11):1519-1533. doi: 10.1261/rna.079233.122. Epub 2022 Aug 30. RNA. 2022. PMID: 36041871 Free PMC article.
-
The C-terminal extension of Lsm4 interacts directly with the 3' end of the histone mRNP and is required for efficient histone mRNA degradation.RNA. 2014 Jan;20(1):88-102. doi: 10.1261/rna.042531.113. Epub 2013 Nov 19. RNA. 2014. PMID: 24255165 Free PMC article.
-
Degradation of oligouridylated histone mRNAs: see UUUUU and goodbye.Wiley Interdiscip Rev RNA. 2014 Jul-Aug;5(4):577-89. doi: 10.1002/wrna.1232. Epub 2014 Apr 1. Wiley Interdiscip Rev RNA. 2014. PMID: 24692427 Review.
-
Expression of metazoan replication-dependent histone genes.Biochimie. 2005 Sep-Oct;87(9-10):827-34. doi: 10.1016/j.biochi.2005.03.012. Epub 2005 Apr 12. Biochimie. 2005. PMID: 16164992 Review.
Cited by
-
5' and 3' modifications controlling RNA degradation: from safeguards to executioners.Philos Trans R Soc Lond B Biol Sci. 2018 Nov 5;373(1762):20180160. doi: 10.1098/rstb.2018.0160. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 30397097 Free PMC article.
-
Terminal nucleotidyl transferases (TENTs) in mammalian RNA metabolism.Philos Trans R Soc Lond B Biol Sci. 2018 Nov 5;373(1762):20180162. doi: 10.1098/rstb.2018.0162. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 30397099 Free PMC article. Review.
-
Uridylation of the histone mRNA stem-loop weakens binding interactions with SLBP while maintaining interactions with 3'hExo.RNA Biol. 2023 Jan;20(1):469-481. doi: 10.1080/15476286.2023.2171760. RNA Biol. 2023. PMID: 37516934 Free PMC article.
-
Cyclin E/CDK2 and feedback from soluble histone protein regulate the S phase burst of histone biosynthesis.Cell Rep. 2023 Jul 25;42(7):112768. doi: 10.1016/j.celrep.2023.112768. Epub 2023 Jul 9. Cell Rep. 2023. PMID: 37428633 Free PMC article.
-
UPF1-Mediated RNA Decay-Danse Macabre in a Cloud.Biomolecules. 2020 Jul 4;10(7):999. doi: 10.3390/biom10070999. Biomolecules. 2020. PMID: 32635561 Free PMC article. Review.
References
-
- Whitfield ML, Zheng L-X, Baldwin A, Ohta T, Hurt MM, Marzluff WF. 2000. Stem-loop binding protein, the protein that binds the 3′ end of histone mRNA, is cell cycle regulated by both translational and posttranslational mechanisms. Mol. Cell. Biol. 20, 4188–4198. (10.1128/MCB.20.12.4188-4198.2000) - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
