

Outer Membrane Vesicles Derived From Escherichia coli Regulate Neutrophil Migration by Induction of Endothelial IL-8
- PMID: 30369908
- PMCID: PMC6194319
- DOI: 10.3389/fmicb.2018.02268
Outer Membrane Vesicles Derived From Escherichia coli Regulate Neutrophil Migration by Induction of Endothelial IL-8
Abstract
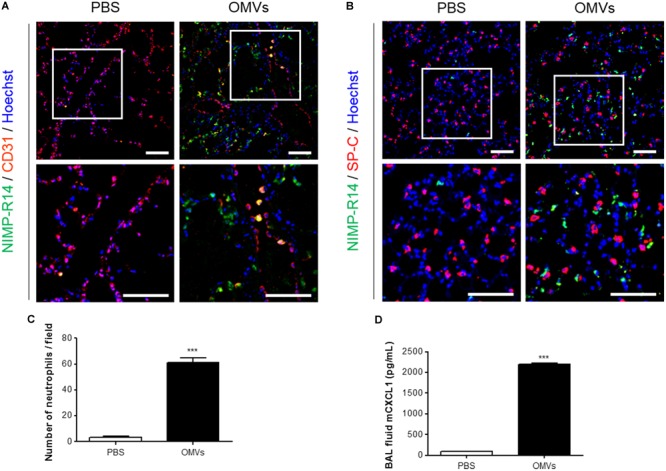
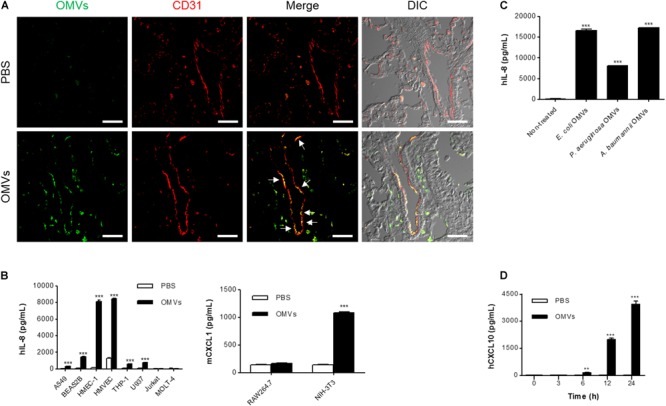
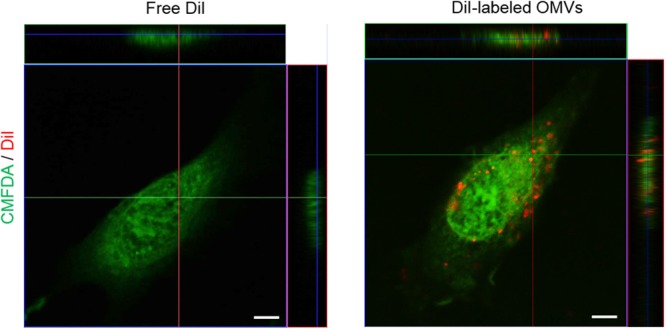
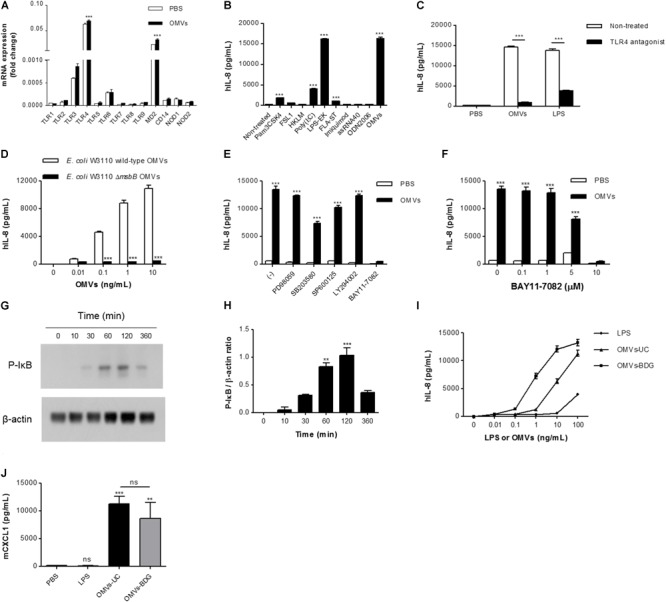
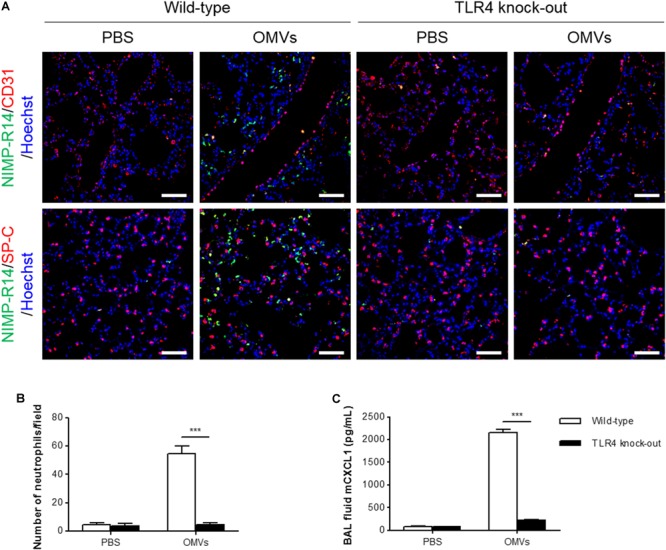
Outer membrane vesicles (OMVs) are spherical, proteolipid nanostructures that are constitutively released by Gram-negative bacteria including Escherichia coli. Although it has been shown that administration of E. coli OMVs stimulates a strong pulmonary inflammatory response with infiltration of neutrophils into the lungs in vivo, the mechanism of E. coli OMV-mediated neutrophil recruitment is poorly characterized. In this study, we observed significant infiltration of neutrophils into the mouse lung tissues in vivo, with increased expression of the neutrophil chemoattractant CXCL1, a murine functional homolog of human IL-8, on intraperitoneal administration of E. coli OMVs. In addition, OMVs and CD31-positive endothelial cells colocalized in the mouse lungs. Moreover, in vitro results showed that E. coli OMVs significantly increased IL-8 release from human microvascular endothelial cells and toll-like receptor (TLR)4 was found to be the main component for recognizing E. coli OMVs among human endothelial cell-associated TLRs. Furthermore, the transmigration of neutrophils was suppressed in the lung tissues obtained from TLR4 knockout mice treated with E. coli OMVs. Taken together, our data demonstrated that E. coli OMVs potently recruit neutrophils into the lung via the release of IL-8/CXCL1 from endothelial cells in TLR4- and NF-κB-dependent manners.
Keywords: IL-8; NF-κB; exosomes; extracellular vesicles; neutrophil; outer membrane vesicles; pulmonary inflammation; toll-like receptor 4.
Figures
Similar articles
-
Outer membrane vesicles derived from Escherichia coli up-regulate expression of endothelial cell adhesion molecules in vitro and in vivo.PLoS One. 2013;8(3):e59276. doi: 10.1371/journal.pone.0059276. Epub 2013 Mar 14. PLoS One. 2013. PMID: 23516621 Free PMC article.
-
Bacterial vesicles block viral replication in macrophages via TLR4-TRIF-axis.Cell Commun Signal. 2023 Mar 28;21(1):65. doi: 10.1186/s12964-023-01086-4. Cell Commun Signal. 2023. PMID: 36978183 Free PMC article.
-
Outer membrane vesicles from pathogenic bacteria initiate an inflammatory response in human endothelial cells.J Surg Res. 2013 Sep;184(1):458-66. doi: 10.1016/j.jss.2013.05.035. Epub 2013 Jun 2. J Surg Res. 2013. PMID: 23800440
-
The role and mechanisms of gram-negative bacterial outer membrane vesicles in inflammatory diseases.Front Immunol. 2023 Jun 16;14:1157813. doi: 10.3389/fimmu.2023.1157813. eCollection 2023. Front Immunol. 2023. PMID: 37398647 Free PMC article. Review.
-
Bacterial outer membrane vesicles as potential biological nanomaterials for antibacterial therapy.Acta Biomater. 2022 Mar 1;140:102-115. doi: 10.1016/j.actbio.2021.12.005. Epub 2021 Dec 9. Acta Biomater. 2022. PMID: 34896632 Review.
Cited by
-
A Review on the Interaction of Acetic Acid Bacteria and Microbes in Food Fermentation: A Microbial Ecology Perspective.Foods. 2024 Aug 14;13(16):2534. doi: 10.3390/foods13162534. Foods. 2024. PMID: 39200461 Free PMC article. Review.
-
Interaction with mammalian enteric viruses alters outer membrane vesicle production and content by commensal bacteria.J Extracell Vesicles. 2022 Jan;11(1):e12172. doi: 10.1002/jev2.12172. J Extracell Vesicles. 2022. PMID: 34981901 Free PMC article.
-
Delivery of Toxins and Effectors by Bacterial Membrane Vesicles.Toxins (Basel). 2021 Nov 26;13(12):845. doi: 10.3390/toxins13120845. Toxins (Basel). 2021. PMID: 34941684 Free PMC article. Review.
-
Bacterial extracellular vesicles isolated from organic dust induce neutrophilic inflammation in the lung.Am J Physiol Lung Cell Mol Physiol. 2020 Dec 1;319(6):L893-L907. doi: 10.1152/ajplung.00107.2020. Epub 2020 Sep 30. Am J Physiol Lung Cell Mol Physiol. 2020. PMID: 32996778 Free PMC article.
-
Released bacterial ATP shapes local and systemic inflammation during abdominal sepsis.Elife. 2024 Aug 20;13:RP96678. doi: 10.7554/eLife.96678. Elife. 2024. PMID: 39163101 Free PMC article.
References
-
- Alaniz R. C., Deatherage B. L., Lara J. C., Cookson B. T. (2007). Membrane vesicles are immunogenic facsimiles of Salmonella typhimurium that potently activate dendritic cells, prime B and T cell responses, and stimulate protective immunity in vivo. J. Immunol. 179 7692–7701. 10.4049/jimmunol.179.11.7692 - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources