

Structure of the membrane-assembled retromer coat determined by cryo-electron tomography
- PMID: 30224749
- PMCID: PMC6173284
- DOI: 10.1038/s41586-018-0526-z
Structure of the membrane-assembled retromer coat determined by cryo-electron tomography
Abstract
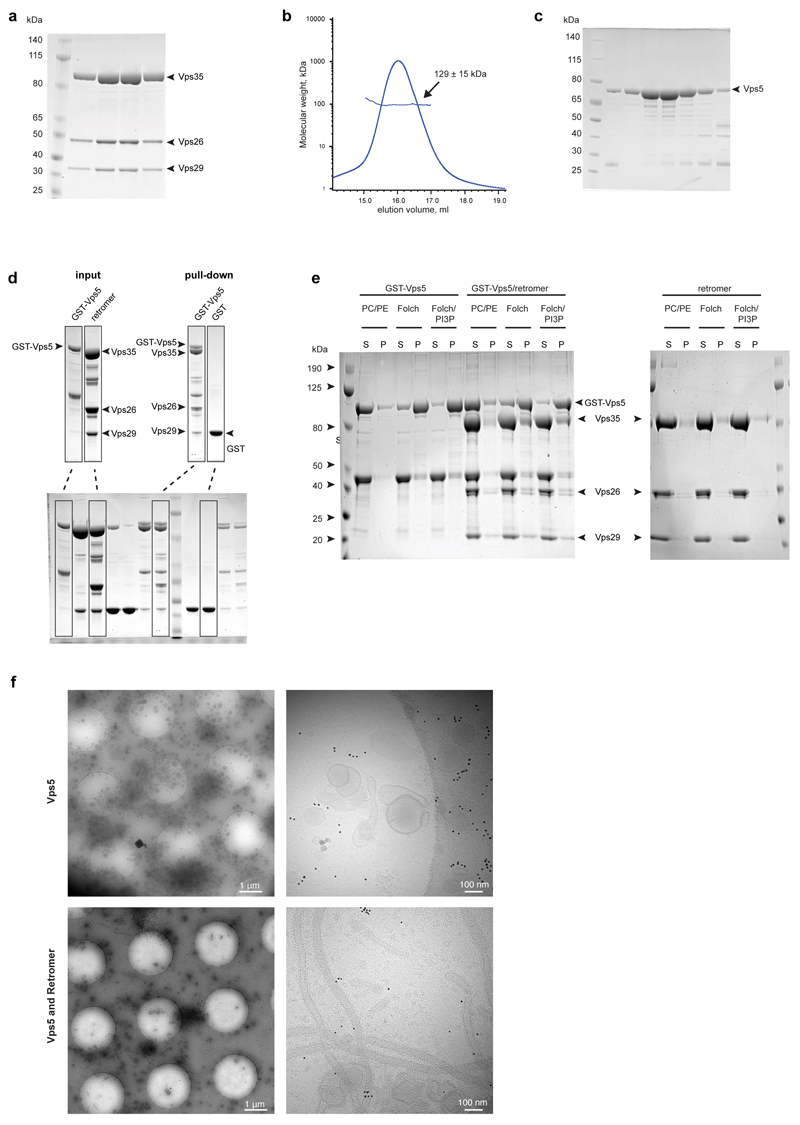
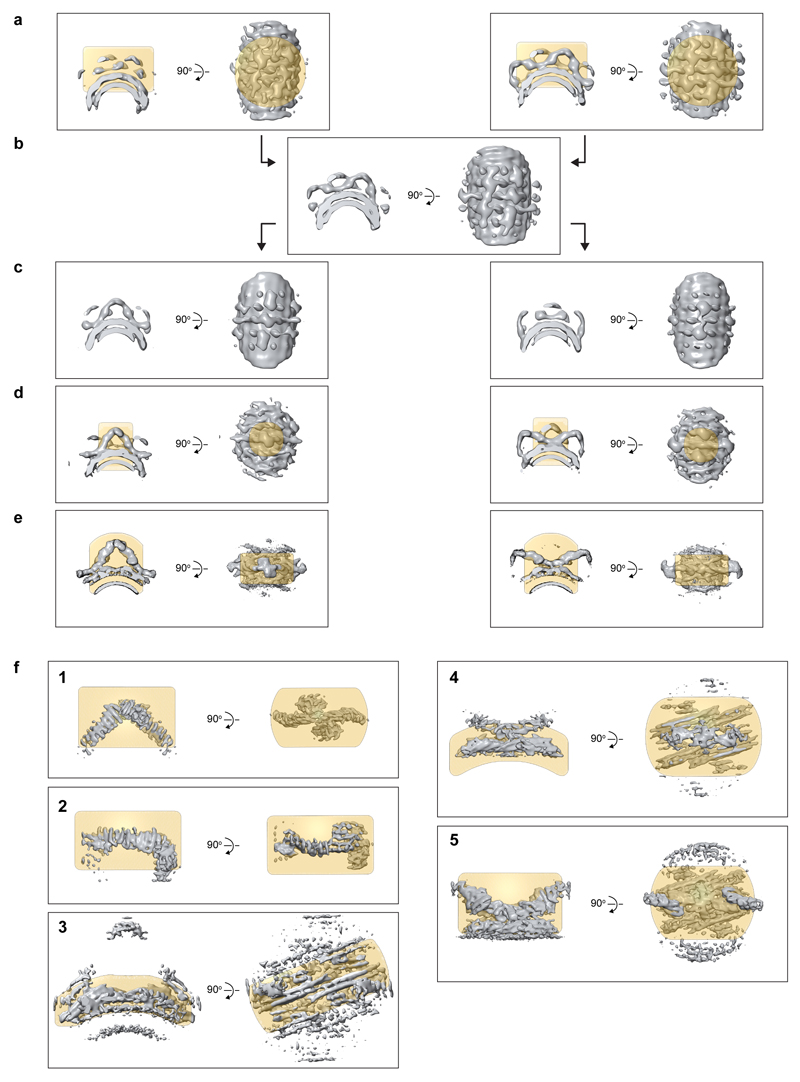
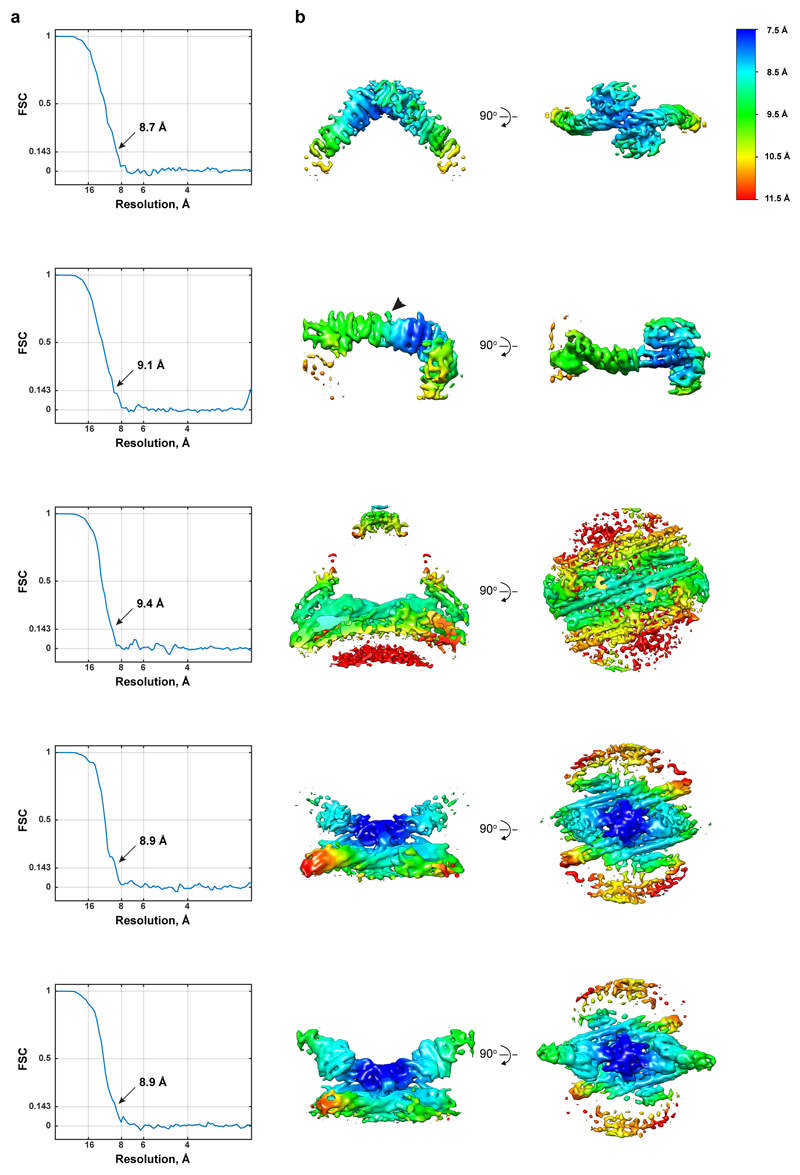
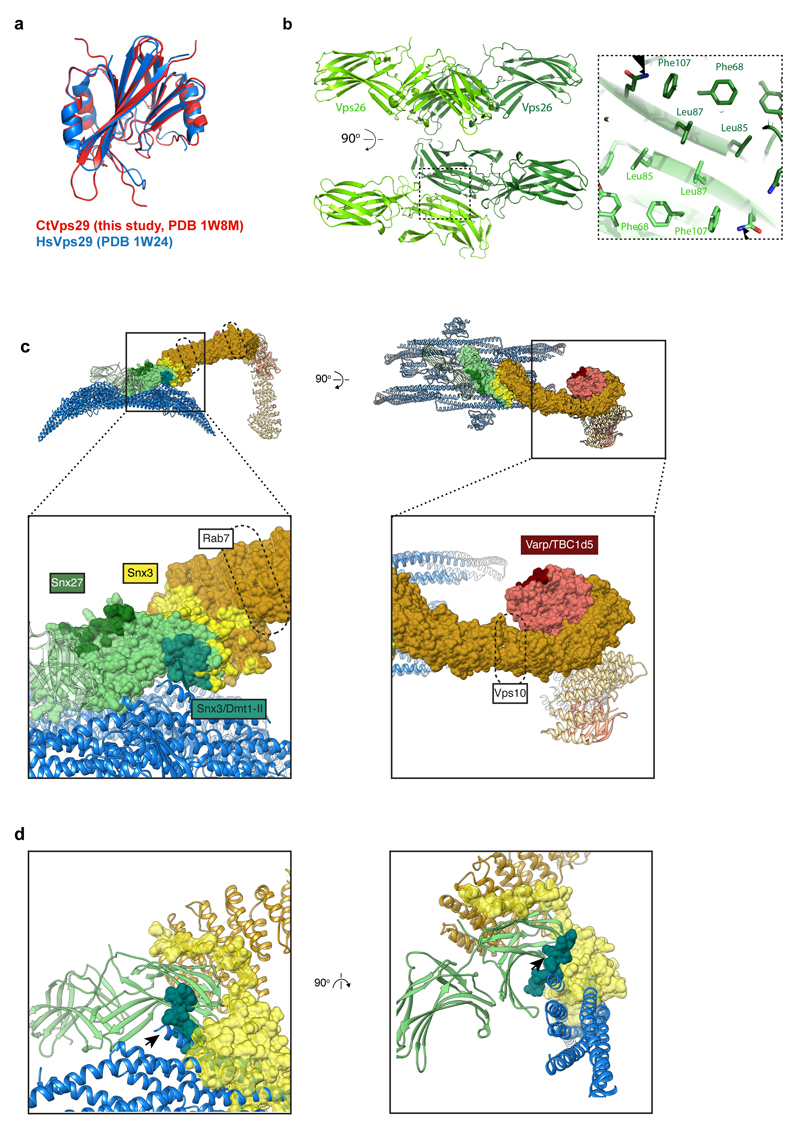
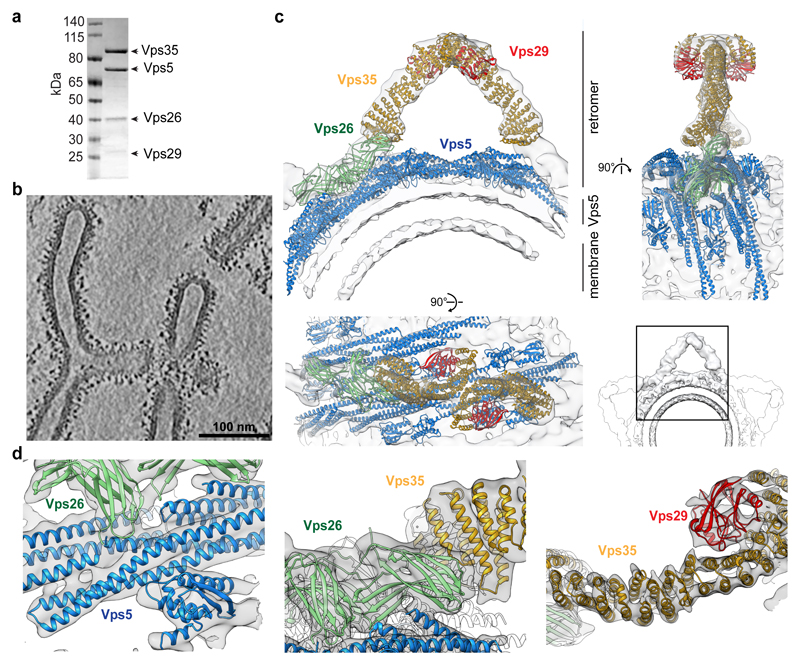
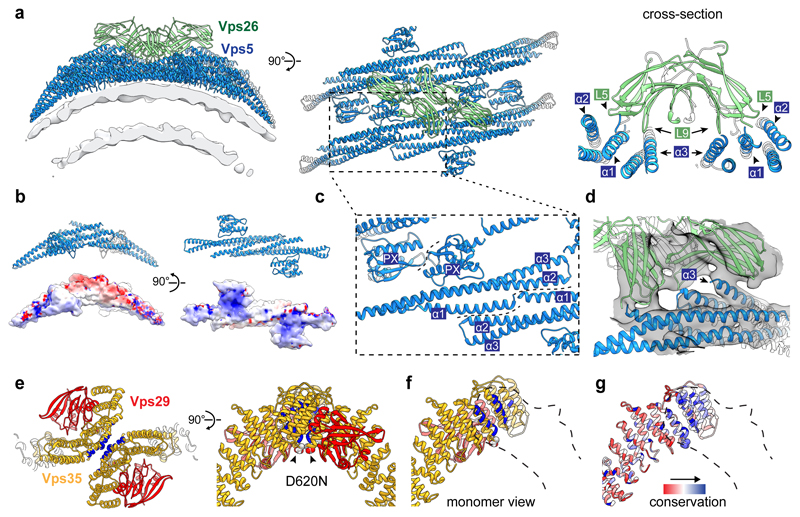
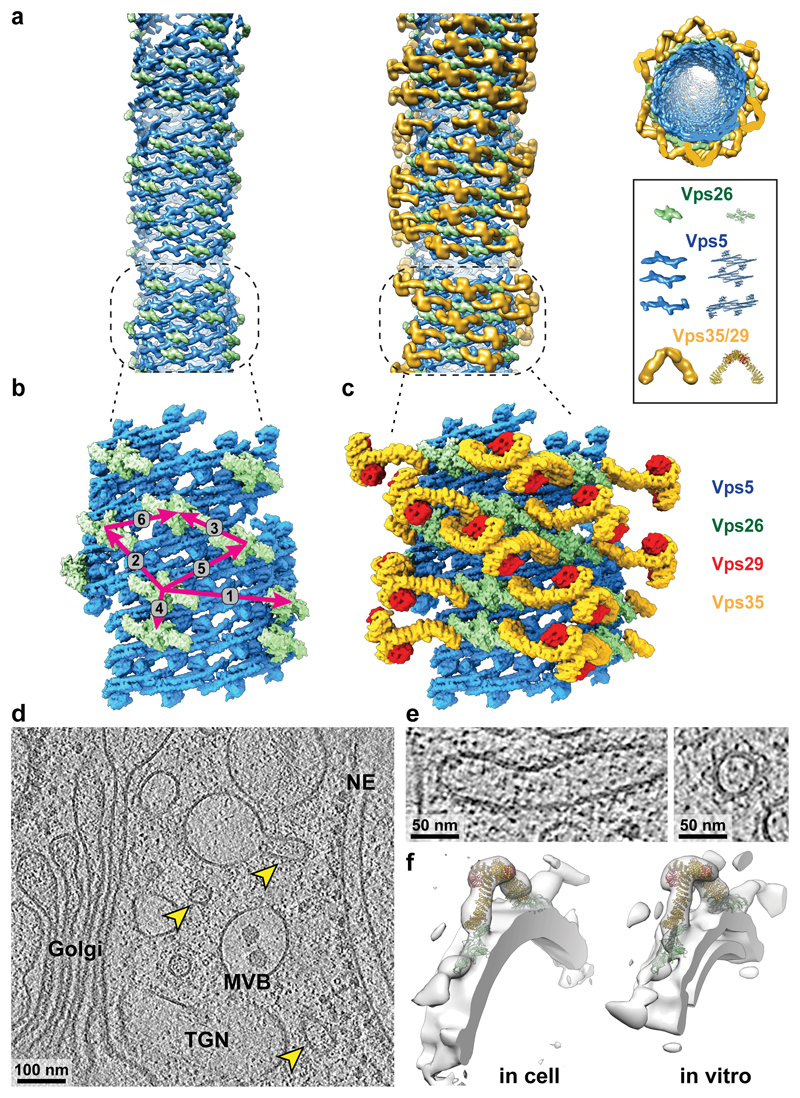
Eukaryotic cells traffic proteins and lipids between different compartments using protein-coated vesicles and tubules. The retromer complex is required to generate cargo-selective tubulovesicular carriers from endosomal membranes1-3. Conserved in eukaryotes, retromer controls the cellular localization and homeostasis of hundreds of transmembrane proteins, and its disruption is associated with major neurodegenerative disorders4-7. How retromer is assembled and how it is recruited to form coated tubules is not known. Here we describe the structure of the retromer complex (Vps26-Vps29-Vps35) assembled on membrane tubules with the bin/amphiphysin/rvs-domain-containing sorting nexin protein Vps5, using cryo-electron tomography and subtomogram averaging. This reveals a membrane-associated Vps5 array, from which arches of retromer extend away from the membrane surface. Vps35 forms the 'legs' of these arches, and Vps29 resides at the apex where it is free to interact with regulatory factors. The bases of the arches connect to each other and to Vps5 through Vps26, and the presence of the same arches on coated tubules within cells confirms their functional importance. Vps5 binds to Vps26 at a position analogous to the previously described cargo- and Snx3-binding site, which suggests the existence of distinct retromer-sorting nexin assemblies. The structure provides insight into the architecture of the coat and its mechanism of assembly, and suggests that retromer promotes tubule formation by directing the distribution of sorting nexin proteins on the membrane surface while providing a scaffold for regulatory-protein interactions.
Conflict of interest statement
The authors declare that they have no competing financial interests
Figures

Comment in
-
Endosomal Sorting: Architecture of the Retromer Coat.Curr Biol. 2018 Dec 3;28(23):R1350-R1352. doi: 10.1016/j.cub.2018.10.040. Curr Biol. 2018. PMID: 30513333
Similar articles
-
Unveiling the cryo-EM structure of retromer.Biochem Soc Trans. 2020 Oct 30;48(5):2261-2272. doi: 10.1042/BST20200552. Biochem Soc Trans. 2020. PMID: 33125482 Free PMC article. Review.
-
Structural Mechanism for Cargo Recognition by the Retromer Complex.Cell. 2016 Dec 1;167(6):1623-1635.e14. doi: 10.1016/j.cell.2016.10.056. Epub 2016 Nov 23. Cell. 2016. PMID: 27889239 Free PMC article.
-
A mechanism for retromer endosomal coat complex assembly with cargo.Proc Natl Acad Sci U S A. 2014 Jan 7;111(1):267-72. doi: 10.1073/pnas.1316482111. Epub 2013 Dec 16. Proc Natl Acad Sci U S A. 2014. PMID: 24344282 Free PMC article.
-
Architecture and mechanism of metazoan retromer:SNX3 tubular coat assembly.Sci Adv. 2021 Mar 24;7(13):eabf8598. doi: 10.1126/sciadv.abf8598. Print 2021 Mar. Sci Adv. 2021. PMID: 33762348 Free PMC article.
-
Updated Insight into the Physiological and Pathological Roles of the Retromer Complex.Int J Mol Sci. 2017 Jul 25;18(8):1601. doi: 10.3390/ijms18081601. Int J Mol Sci. 2017. PMID: 28757549 Free PMC article. Review.
Cited by
-
PI4P and BLOC-1 remodel endosomal membranes into tubules.J Cell Biol. 2022 Nov 7;221(11):e202110132. doi: 10.1083/jcb.202110132. Epub 2022 Sep 28. J Cell Biol. 2022. PMID: 36169638 Free PMC article.
-
Visualizing membrane trafficking through the electron microscope: cryo-tomography of coat complexes.Acta Crystallogr D Struct Biol. 2019 May 1;75(Pt 5):467-474. doi: 10.1107/S2059798319005011. Epub 2019 Apr 30. Acta Crystallogr D Struct Biol. 2019. PMID: 31063149 Free PMC article.
-
Phosphoinositides in plant-pathogen interaction: trends and perspectives.Stress Biol. 2023 Apr 6;3(1):4. doi: 10.1007/s44154-023-00082-5. Stress Biol. 2023. PMID: 37676371 Free PMC article. Review.
-
Protein-membrane interactions: sensing and generating curvature.Trends Biochem Sci. 2024 May;49(5):401-416. doi: 10.1016/j.tibs.2024.02.005. Epub 2024 Mar 19. Trends Biochem Sci. 2024. PMID: 38508884 Review.
-
Unveiling the cryo-EM structure of retromer.Biochem Soc Trans. 2020 Oct 30;48(5):2261-2272. doi: 10.1042/BST20200552. Biochem Soc Trans. 2020. PMID: 33125482 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
