

Phenotypic and Functional Signatures of Herpes Simplex Virus-Specific Effector Memory CD73+CD45RAhighCCR7lowCD8+ TEMRA and CD73+CD45RAlowCCR7lowCD8+ TEM Cells Are Associated with Asymptomatic Ocular Herpes
- PMID: 30201808
- PMCID: PMC6195422
- DOI: 10.4049/jimmunol.1800725
Phenotypic and Functional Signatures of Herpes Simplex Virus-Specific Effector Memory CD73+CD45RAhighCCR7lowCD8+ TEMRA and CD73+CD45RAlowCCR7lowCD8+ TEM Cells Are Associated with Asymptomatic Ocular Herpes
Abstract
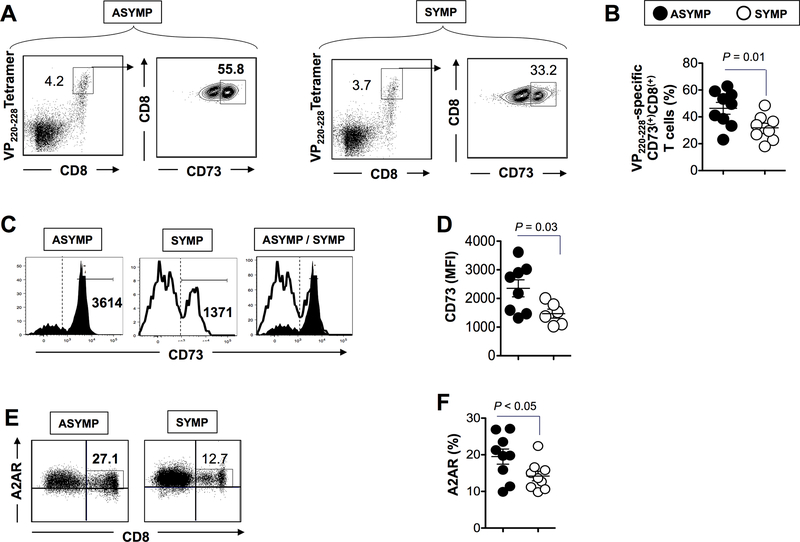
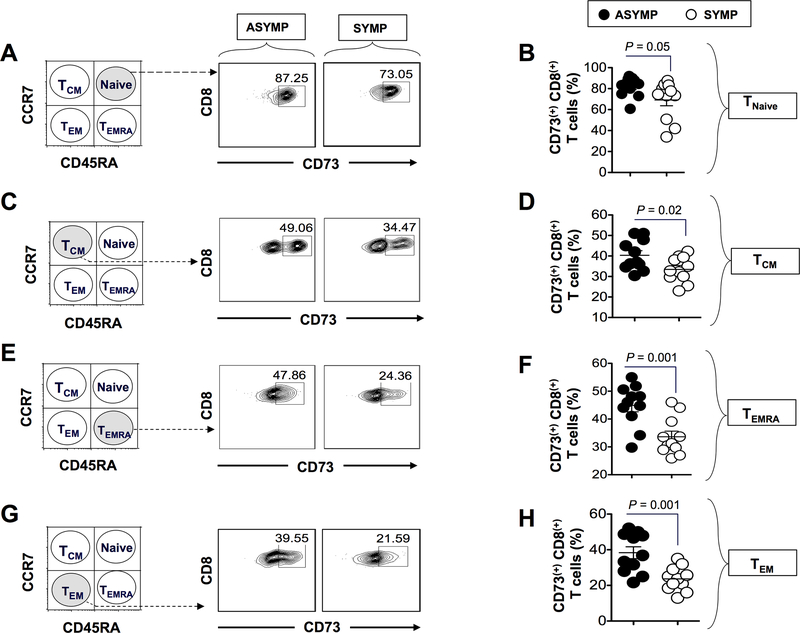
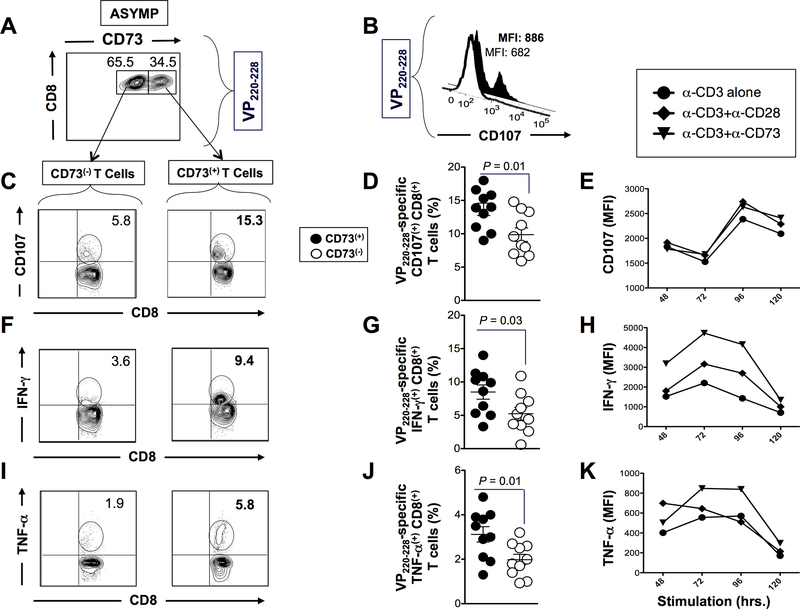
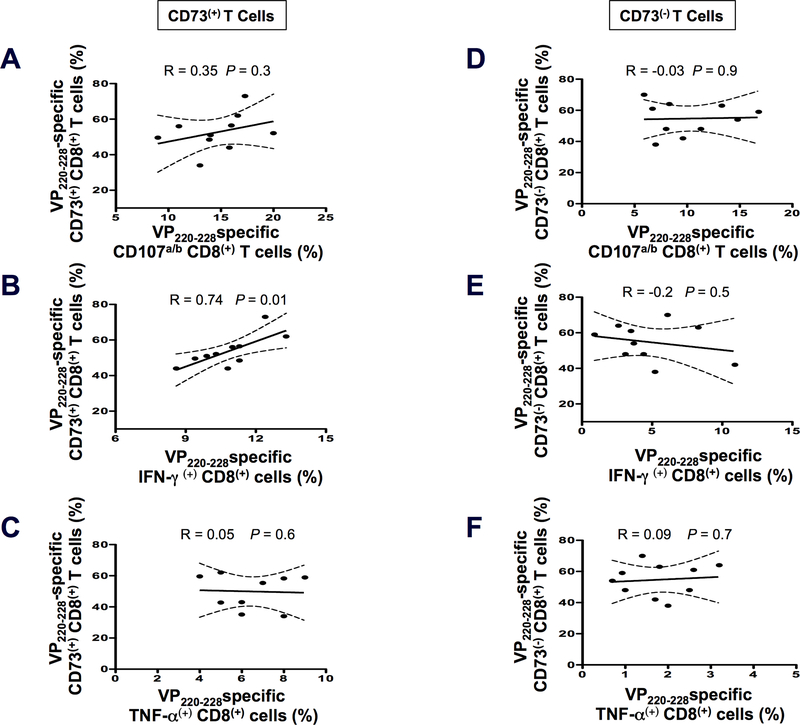
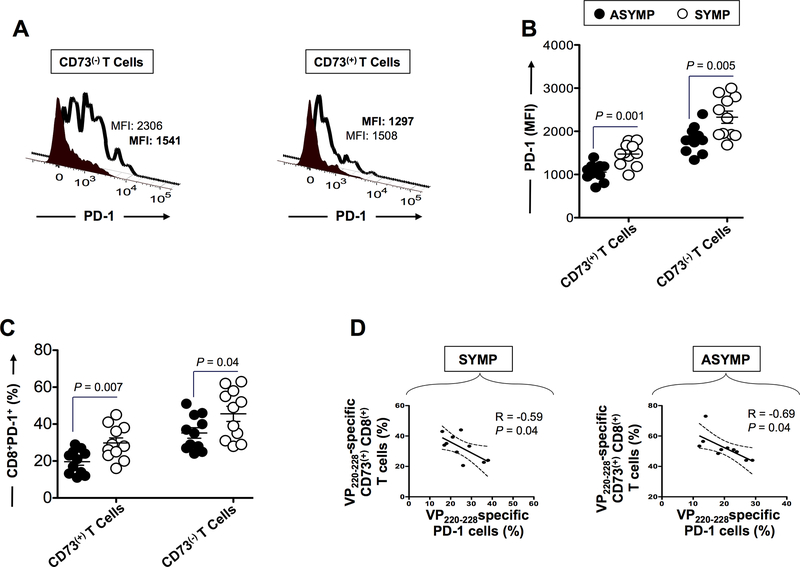
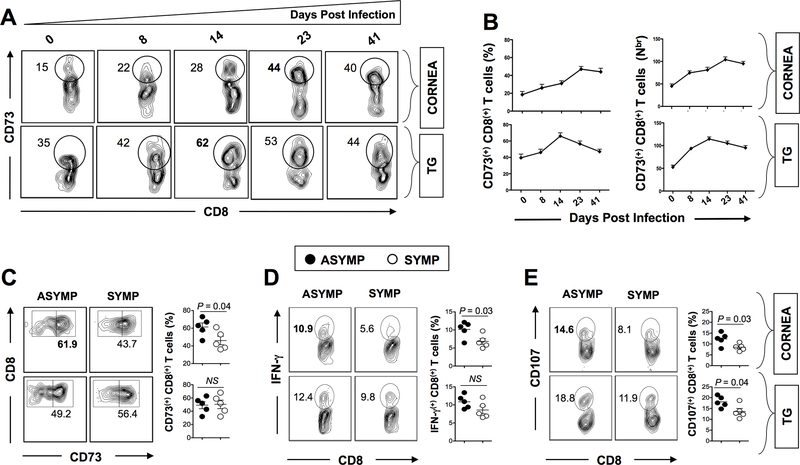
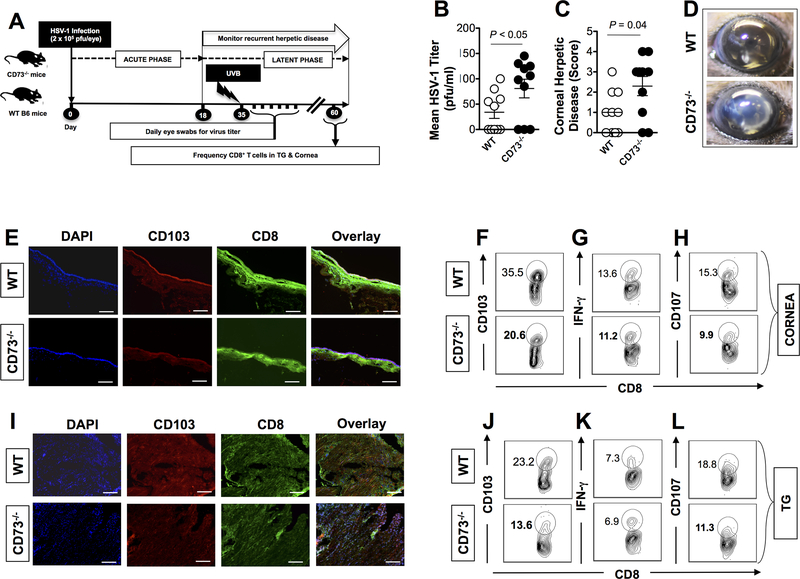
HSV type 1 (HSV-1)-specific CD8+ T cells protect from herpes infection and disease. However, the nature of protective CD8+ T cells in HSV-1 seropositive healthy asymptomatic (ASYMP) individuals (with no history of clinical herpes disease) remains to be determined. In this study, we compared the phenotype and function of HSV-specific CD8+ T cells from HLA-A*02:01-positive ASYMP and symptomatic (SYMP) individuals (with a documented history of numerous episodes of recurrent ocular herpetic disease). We report that although SYMP and ASYMP individuals have similar frequencies of HSV-specific CD8+ T cells, the "naturally" protected ASYMP individuals have a significantly higher proportion of multifunctional HSV-specific effector memory CD8+ T cells (CD73+CD45RAhighCCR7lowCD8+ effector memory RA (TEMRA) and CD73+CD45RAlowCCR7lowCD8+ effector memory (TEM) as compared with SYMP individuals. Similar to humans, HSV-1-infected ASYMP B6 mice had frequent multifunctional HSV-specific CD73+CD8+ T cells in the cornea, as compared with SYMP mice. Moreover, in contrast to wild type B6, CD73-/- deficient mice infected ocularly with HSV-1 developed more recurrent corneal herpetic infection and disease. This was associated with less functional CD8+ T cells in the cornea and trigeminal ganglia, the sites of acute and latent infection. The phenotypic and functional characteristics of HSV-specific circulating and in situ CD73+CD8+ T cells, demonstrated in both ASYMP humans and mice, suggest a positive role for effector memory CD8+ T cells expressing the CD73 costimulatory molecule in the protection against ocular herpes infection and disease. These findings are important for the development of safe and effective T cell-based herpes immunotherapy.
Copyright © 2018 by The American Association of Immunologists, Inc.
Conflict of interest statement
Conflict of Interest: The authors have declared that no conflicts of interest exist.
Figures
Similar articles
-
Phenotypic and functional characterization of herpes simplex virus glycoprotein B epitope-specific effector and memory CD8+ T cells from symptomatic and asymptomatic individuals with ocular herpes.J Virol. 2015 Apr;89(7):3776-92. doi: 10.1128/JVI.03419-14. Epub 2015 Jan 21. J Virol. 2015. PMID: 25609800 Free PMC article.
-
Human Asymptomatic Epitopes Identified from the Herpes Simplex Virus Tegument Protein VP13/14 (UL47) Preferentially Recall Polyfunctional Effector Memory CD44high CD62Llow CD8+ TEM Cells and Protect Humanized HLA-A*02:01 Transgenic Mice against Ocular Herpesvirus Infection.J Virol. 2017 Jan 3;91(2):e01793-16. doi: 10.1128/JVI.01793-16. Print 2017 Jan 15. J Virol. 2017. PMID: 27847359 Free PMC article.
-
Unique Type I Interferon, Expansion/Survival Cytokines, and JAK/STAT Gene Signatures of Multifunctional Herpes Simplex Virus-Specific Effector Memory CD8+ TEM Cells Are Associated with Asymptomatic Herpes in Humans.J Virol. 2019 Feb 5;93(4):e01882-18. doi: 10.1128/JVI.01882-18. Print 2019 Feb 15. J Virol. 2019. PMID: 30487281 Free PMC article.
-
Combinatorial Herpes Simplex Vaccine Strategies: From Bedside to Bench and Back.Front Immunol. 2022 Apr 25;13:849515. doi: 10.3389/fimmu.2022.849515. eCollection 2022. Front Immunol. 2022. PMID: 35547736 Free PMC article. Review.
-
Asymptomatic memory CD8+ T cells: from development and regulation to consideration for human vaccines and immunotherapeutics.Hum Vaccin Immunother. 2014;10(4):945-63. doi: 10.4161/hv.27762. Epub 2014 Feb 5. Hum Vaccin Immunother. 2014. PMID: 24499824 Free PMC article. Review.
Cited by
-
Upregulation of Multiple CD8+ T Cell Exhaustion Pathways Is Associated with Recurrent Ocular Herpes Simplex Virus Type 1 Infection.J Immunol. 2020 Jul 15;205(2):454-468. doi: 10.4049/jimmunol.2000131. Epub 2020 Jun 15. J Immunol. 2020. PMID: 32540992 Free PMC article.
-
Therapeutic prime/pull vaccination of HSV-2-infected guinea pigs with the ribonucleotide reductase 2 (RR2) protein and CXCL11 chemokine boosts antiviral local tissue-resident and effector memory CD4+ and CD8+ T cells and protects against recurrent genital herpes.J Virol. 2024 May 14;98(5):e0159623. doi: 10.1128/jvi.01596-23. Epub 2024 Apr 8. J Virol. 2024. PMID: 38587378 Free PMC article.
-
Therapeutic Mucosal Vaccination of Herpes Simplex Virus 2-Infected Guinea Pigs with Ribonucleotide Reductase 2 (RR2) Protein Boosts Antiviral Neutralizing Antibodies and Local Tissue-Resident CD4+ and CD8+ TRM Cells Associated with Protection against Recurrent Genital Herpes.J Virol. 2019 Apr 17;93(9):e02309-18. doi: 10.1128/JVI.02309-18. Print 2019 May 1. J Virol. 2019. PMID: 30787156 Free PMC article.
-
Distinct transcriptomic and epigenomic modalities underpin human memory T cell subsets and their activation potential.Commun Biol. 2023 Apr 3;6(1):363. doi: 10.1038/s42003-023-04747-9. Commun Biol. 2023. PMID: 37012418 Free PMC article.
-
Intramuscular Vaccination With the HSV-1(VC2) Live-Attenuated Vaccine Strain Confers Protection Against Viral Ocular Immunopathogenesis Associated With γδT Cell Intracorneal Infiltration.Front Immunol. 2021 Nov 15;12:789454. doi: 10.3389/fimmu.2021.789454. eCollection 2021. Front Immunol. 2021. PMID: 34868077 Free PMC article.
References
-
- Dervillez X, Qureshi H, Chentoufi AA, Khan AA, Kritzer E, Yu DC, Diaz OR, Gottimukkala C, Kalantari M, Villacres MC, Scarfone VM, McKinney DM, Sidney J, Sette A, Nesburn AB, Wechsler SL, and BenMohamed L. 2013. Asymptomatic HLA-A*02:01-restricted epitopes from herpes simplex virus glycoprotein B preferentially recall polyfunctional CD8+ T cells from seropositive asymptomatic individuals and protect HLA transgenic mice against ocular herpes. J Immunol 191: 5124–5138. - PMC - PubMed
-
- Chentoufi AA, Zhang X, Lamberth K, Dasgupta G, Bettahi I, Nguyen A, Wu M, Zhu X, Mohebbi A, Buus S, Wechsler SL, Nesburn AB, and BenMohamed L. 2008. HLA-A*0201-restricted CD8+ cytotoxic T lymphocyte epitopes identified from herpes simplex virus glycoprotein D. J Immunol 180: 426–437. - PubMed
-
- Zhang X, Dervillez X, Chentoufi AA, Badakhshan T, Bettahi I, and BenMohamed L. 2012. Targeting the genital tract mucosa with a lipopeptide/recombinant adenovirus prime/boost vaccine induces potent and long-lasting CD8+ T cell immunity against herpes: importance of MyD88. J Immunol 189: 4496–4509. - PMC - PubMed
-
- Samandary S, Kridane-Miledi H, Sandoval JS, Choudhury Z, Langa-Vives F, Spencer D, Chentoufi AA, Lemonnier FA, and BenMohamed L. 2014. Associations of HLA-A, HLA-B and HLA-C alleles frequency with prevalence of herpes simplex virus infections and diseases across global populations: implication for the development of an universal CD8+ T-cell epitope-based vaccine. Hum Immunol 75: 715–729. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials