

Roles of microRNAs in mammalian reproduction: from the commitment of germ cells to peri-implantation embryos
- PMID: 30151880
- PMCID: PMC7379200
- DOI: 10.1111/brv.12459
Roles of microRNAs in mammalian reproduction: from the commitment of germ cells to peri-implantation embryos
Abstract
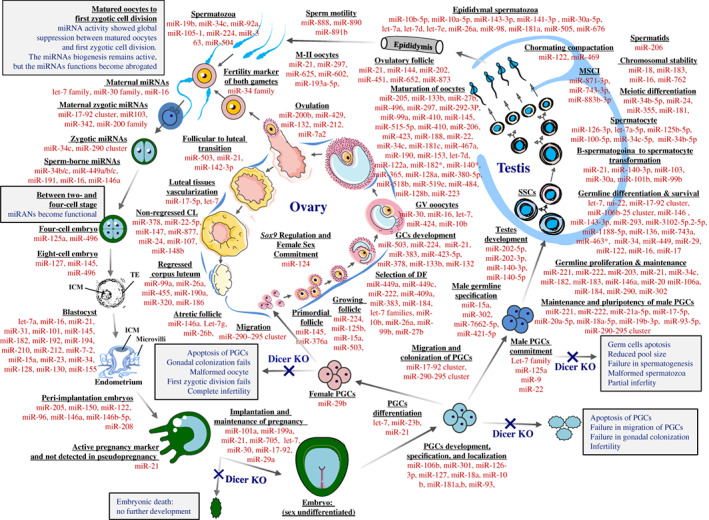
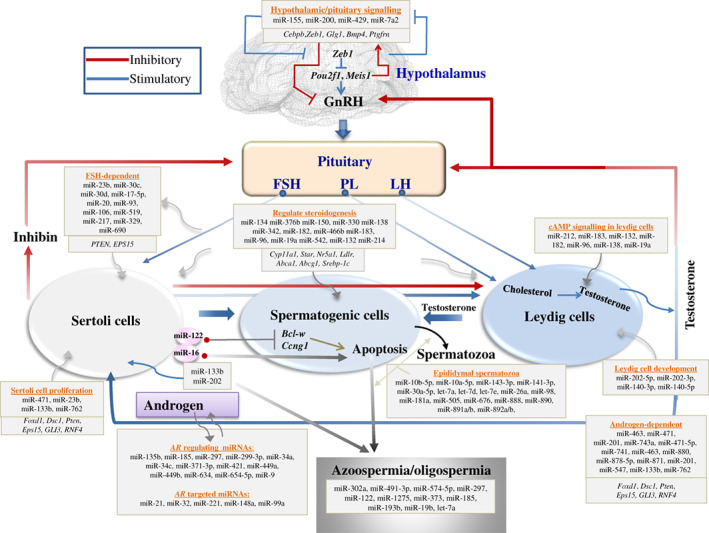
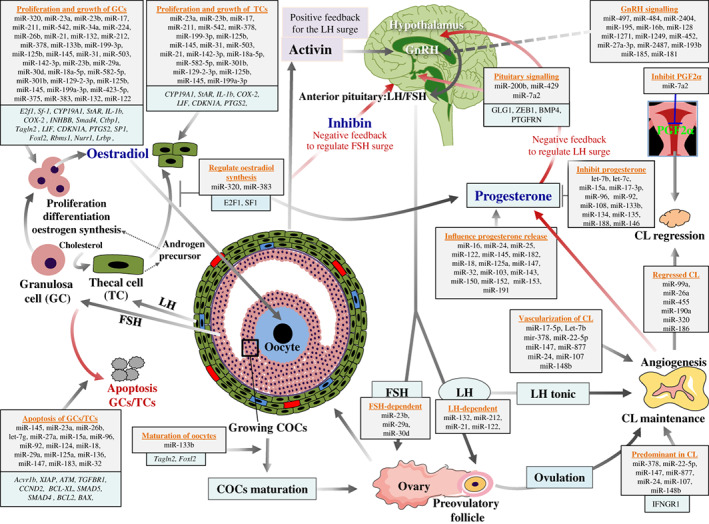
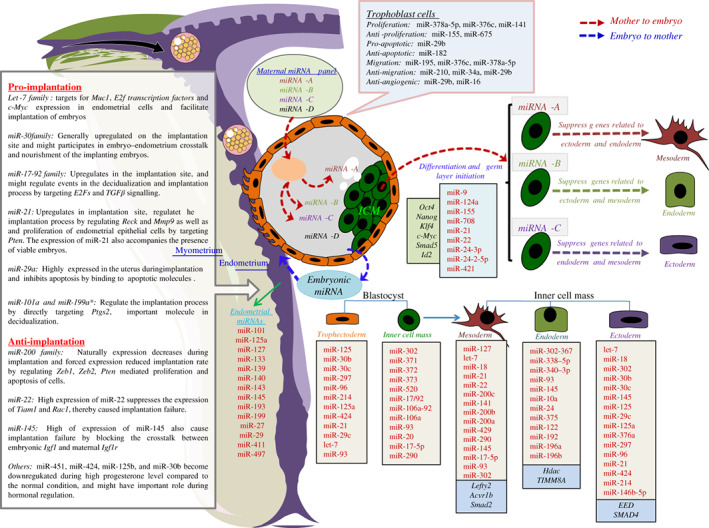
MicroRNAs (miRNAs) are active regulators of numerous biological and physiological processes including most of the events of mammalian reproduction. Understanding the biological functions of miRNAs in the context of mammalian reproduction will allow a better and comparative understanding of fertility and sterility in male and female mammals. Herein, we summarize recent progress in miRNA-mediated regulation of mammalian reproduction and highlight the significance of miRNAs in different aspects of mammalian reproduction including the biogenesis of germ cells, the functionality of reproductive organs, and the development of early embryos. Furthermore, we focus on the gene expression regulatory feedback loops involving hormones and miRNA expression to increase our understanding of germ cell commitment and the functioning of reproductive organs. Finally, we discuss the influence of miRNAs on male and female reproductive failure, and provide perspectives for future studies on this topic.
Keywords: gametogenesis; germ layer specification; hormonal balance; mammalian reproduction; miRNAs; peri-implantation.
© 2018 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures

Similar articles
-
Functions and mechanism of noncoding RNA in regulation and differentiation of male mammalian reproduction.Cell Biochem Funct. 2023 Oct;41(7):767-778. doi: 10.1002/cbf.3838. Epub 2023 Aug 16. Cell Biochem Funct. 2023. PMID: 37583312 Review.
-
MiRNAs in the Peri-Implantation Period: Contribution to Embryo-Maternal Communication in Pigs.Int J Mol Sci. 2020 Mar 23;21(6):2229. doi: 10.3390/ijms21062229. Int J Mol Sci. 2020. PMID: 32210170 Free PMC article. Review.
-
The Role of MicroRNAs in Mammalian Fertility: From Gametogenesis to Embryo Implantation.Int J Mol Sci. 2020 Jan 16;21(2):585. doi: 10.3390/ijms21020585. Int J Mol Sci. 2020. PMID: 31963271 Free PMC article. Review.
-
Review: MicroRNAs in assisted reproduction and their potential role in IVF failure.In Vivo. 2015 Mar-Apr;29(2):169-75. In Vivo. 2015. PMID: 25792643 Review.
-
Recent advances in mammalian reproductive biology.Sci China Life Sci. 2020 Jan;63(1):18-58. doi: 10.1007/s11427-019-1572-7. Epub 2019 Nov 29. Sci China Life Sci. 2020. PMID: 31813094 Review.
Cited by
-
microRNAs Mediated Regulation of the Ribosomal Proteins and its Consequences on the Global Translation of Proteins.Cells. 2021 Jan 8;10(1):110. doi: 10.3390/cells10010110. Cells. 2021. PMID: 33435549 Free PMC article. Review.
-
Evolution after Whole-Genome Duplication: Teleost MicroRNAs.Mol Biol Evol. 2021 Jul 29;38(8):3308-3331. doi: 10.1093/molbev/msab105. Mol Biol Evol. 2021. PMID: 33871629 Free PMC article.
-
The role of miRNAs in regulating adrenal and gonadal steroidogenesis.J Mol Endocrinol. 2020 Jan;64(1):R21-R43. doi: 10.1530/JME-19-0105. J Mol Endocrinol. 2020. PMID: 31671401 Free PMC article. Review.
-
Answer to Controversy: miR-10a Replacement Approaches Do Not Offer Protection against Chemotherapy-Induced Gonadotoxicity in Mouse Model.Int J Mol Sci. 2019 Oct 8;20(19):4958. doi: 10.3390/ijms20194958. Int J Mol Sci. 2019. PMID: 31597292 Free PMC article.
-
Sperm miR-142-3p Reprogramming Mediates Paternal Pre-Pregnancy Caffeine Exposure-Induced Non-Alcoholic Steatohepatitis in Male Offspring Rats.Adv Sci (Weinh). 2024 Nov;11(42):e2405592. doi: 10.1002/advs.202405592. Epub 2024 Sep 18. Adv Sci (Weinh). 2024. PMID: 39291441 Free PMC article.
References
-
- Abd El Naby, W. S. , Hagos, T. H. , Hossain, M. M. , Salilew‐Wondim, D. , Gad, A. Y. , Rings, F. , Cinar, M. U. , Tholen, E. , Looft, C. , Schellander, K. , Hoelker, M. & Tesfaye, D. (2013). Expression analysis of regulatory microRNAs in bovine cumulus oocyte complex and preimplantation embryos. Zygote 21, 31–51. - PubMed
-
- Abu‐Halima, M. , Hammadeh, M. , Backes, C. , Fischer, U. , Leidinger, P. , Lubbad, A. M. , Keller, A. & Meese, E. (2014). Panel of five microRNAs as potential biomarkers for the diagnosis and assessment of male infertility. Fertility and Sterility 102, 989–997.e1. - PubMed
-
- Alford, C. , Toloubeydokhti, T. , Al‐Katanani, Y. , Drury, K. C. , Williams, R. & Chenini, N. (2007). The expression of microRNA (miRNA) mir‐23a and 23b and their target gene, CYP19A1 (aromatase) in follicular cells obtained from women undergoing ART. Fertility and Sterility 88, S166–S167.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
