

Caspase-11 auto-proteolysis is crucial for noncanonical inflammasome activation
- PMID: 30135078
- PMCID: PMC6122968
- DOI: 10.1084/jem.20180589
Caspase-11 auto-proteolysis is crucial for noncanonical inflammasome activation
Abstract
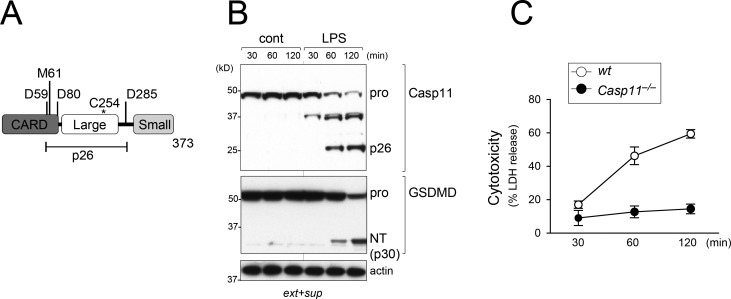
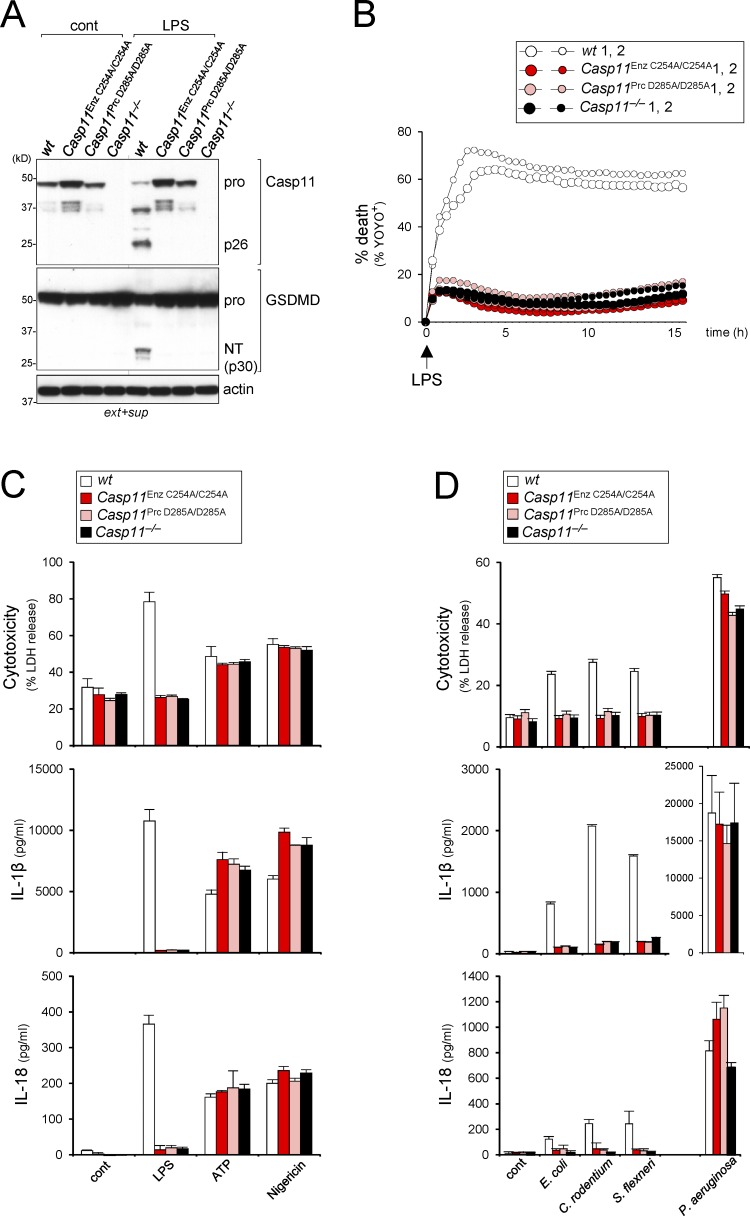
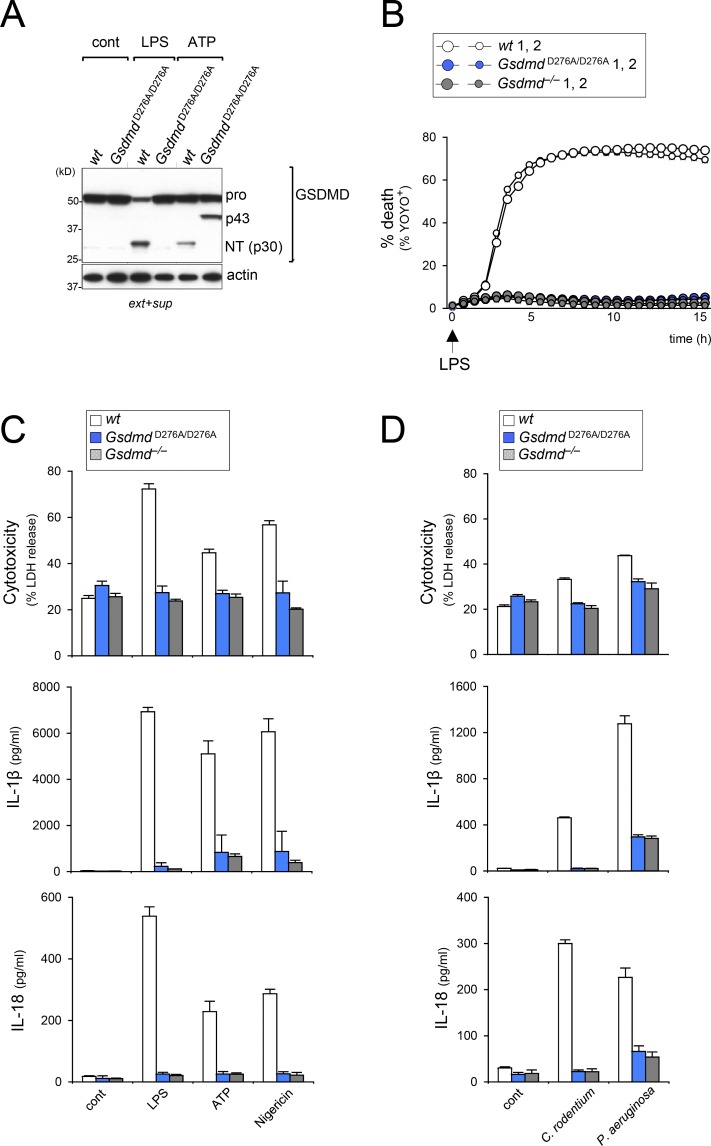
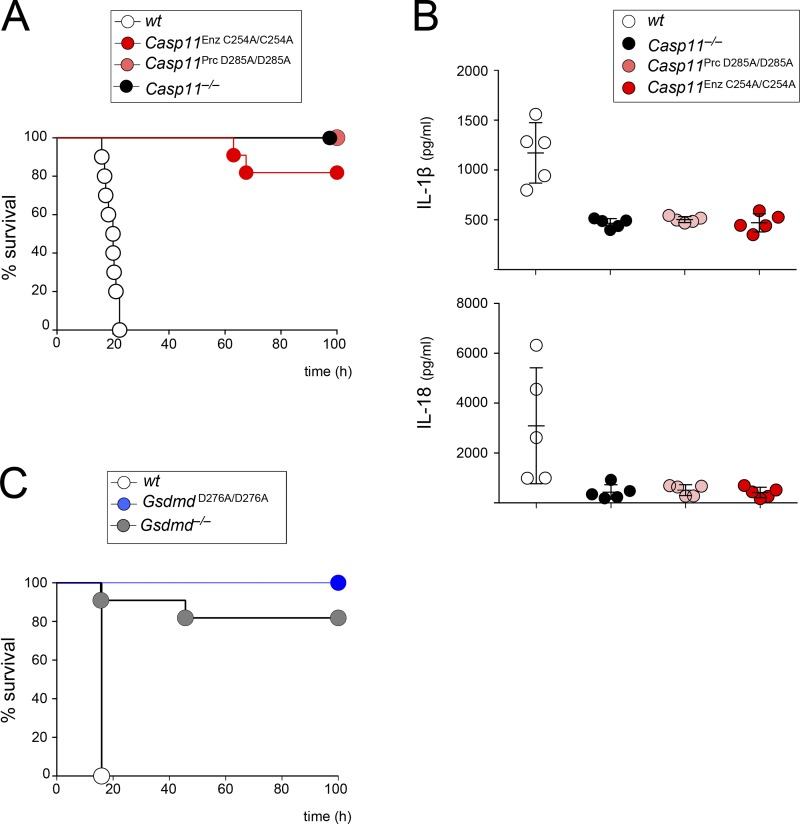
Intracellular LPS sensing by caspase-4/5/11 triggers proteolytic activation of pore-forming gasdermin D (GSDMD), leading to pyroptotic cell death in Gram-negative bacteria-infected cells. Involvement of caspase-4/5/11 and GSDMD in inflammatory responses, such as lethal sepsis, makes them highly desirable drug targets. Using knock-in (KI) mouse strains, we herein provide genetic evidence to show that caspase-11 auto-cleavage at the inter-subunit linker is essential for optimal catalytic activity and subsequent proteolytic cleavage of GSDMD. Macrophages from caspase-11-processing dead KI mice (Casp11Prc D285A/D285A ) exhibit defective caspase-11 auto-processing and phenocopy Casp11-/- and caspase-11 enzymatically dead KI (Casp11Enz C254A/C254A ) macrophages in attenuating responses to cytoplasmic LPS or Gram-negative bacteria infection. GsdmdD276A/D276A KI macrophages also fail to cleave GSDMD and are hypo-responsive to inflammasome stimuli, confirming that the GSDMD Asp276 residue is a nonredundant and indispensable site for proteolytic activation of GSDMD. Our data highlight the role of caspase-11 self-cleavage as a critical regulatory step for GSDMD processing and response against Gram-negative bacteria.
© 2018 Genentech.
Figures
Similar articles
-
Noncanonical inflammasome signaling elicits gasdermin D-dependent neutrophil extracellular traps.Sci Immunol. 2018 Aug 24;3(26):eaar6676. doi: 10.1126/sciimmunol.aar6676. Sci Immunol. 2018. PMID: 30143554
-
Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling.Nature. 2015 Oct 29;526(7575):666-71. doi: 10.1038/nature15541. Epub 2015 Sep 16. Nature. 2015. PMID: 26375259
-
Gasdermin D Promotes AIM2 Inflammasome Activation and Is Required for Host Protection against Francisella novicida.J Immunol. 2018 Dec 15;201(12):3662-3668. doi: 10.4049/jimmunol.1800788. Epub 2018 Nov 7. J Immunol. 2018. PMID: 30404813 Free PMC article.
-
Emerging Insights into Noncanonical Inflammasome Recognition of Microbes.J Mol Biol. 2018 Jan 19;430(2):207-216. doi: 10.1016/j.jmb.2017.10.003. Epub 2017 Oct 7. J Mol Biol. 2018. PMID: 29017836 Free PMC article. Review.
-
Caspase-11 non-canonical inflammasome: a critical sensor of intracellular lipopolysaccharide in macrophage-mediated inflammatory responses.Immunology. 2017 Oct;152(2):207-217. doi: 10.1111/imm.12787. Epub 2017 Jul 31. Immunology. 2017. PMID: 28695629 Free PMC article. Review.
Cited by
-
Recent Insights on Inflammasomes, Gasdermin Pores, and Pyroptosis.Cold Spring Harb Perspect Biol. 2020 May 1;12(5):a036392. doi: 10.1101/cshperspect.a036392. Cold Spring Harb Perspect Biol. 2020. PMID: 31570336 Free PMC article. Review.
-
Inflammasomes: Their Role in Normal and Complicated Pregnancies.J Immunol. 2019 Dec 1;203(11):2757-2769. doi: 10.4049/jimmunol.1900901. J Immunol. 2019. PMID: 31740550 Free PMC article. Review.
-
Inflammatory caspase substrate specificities.mBio. 2024 Jul 17;15(7):e0297523. doi: 10.1128/mbio.02975-23. Epub 2024 Jun 5. mBio. 2024. PMID: 38837391 Free PMC article. Review.
-
The Arg/N-Degron Pathway-A Potential Running Back in Fine-Tuning the Inflammatory Response?Biomolecules. 2020 Jun 14;10(6):903. doi: 10.3390/biom10060903. Biomolecules. 2020. PMID: 32545869 Free PMC article.
-
Intracellular innate immune receptors: Life inside the cell.Immunol Rev. 2020 Sep;297(1):5-12. doi: 10.1111/imr.12912. Immunol Rev. 2020. PMID: 32856334 Free PMC article. No abstract available.
References
-
- Biasini M., Bienert S., Waterhouse A., Arnold K., Studer G., Schmidt T., Kiefer F., Gallo Cassarino T., Bertoni M., Bordoli L., and Schwede T.. 2014. SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Res. 42(W1):W252-258. 10.1093/nar/gku340 - DOI - PMC - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
