

Unified mechanisms for self-RNA recognition by RIG-I Singleton-Merten syndrome variants
- PMID: 30047865
- PMCID: PMC6086658
- DOI: 10.7554/eLife.38958
Unified mechanisms for self-RNA recognition by RIG-I Singleton-Merten syndrome variants
Abstract
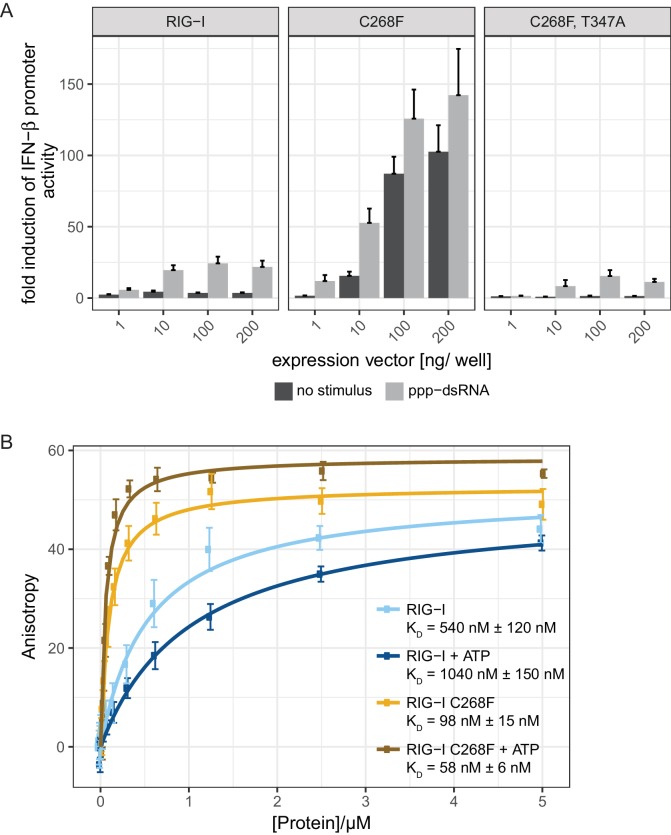
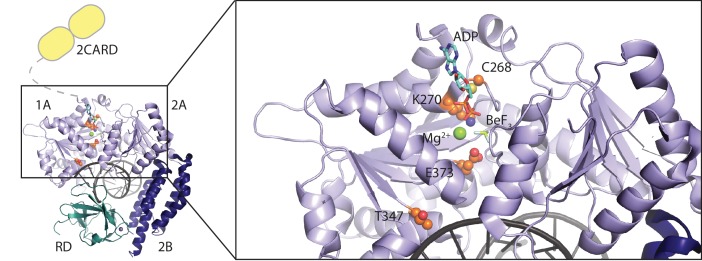
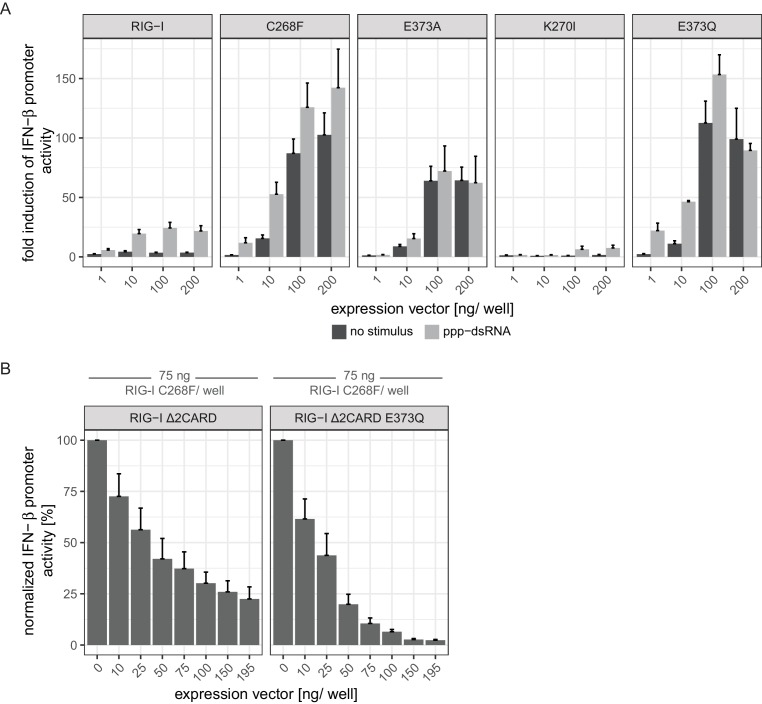
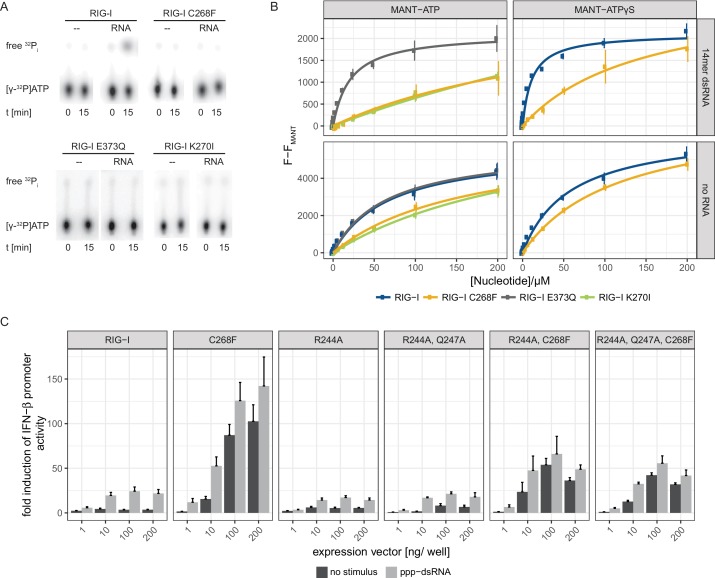
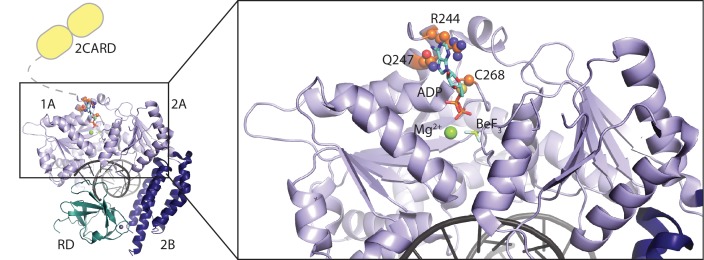
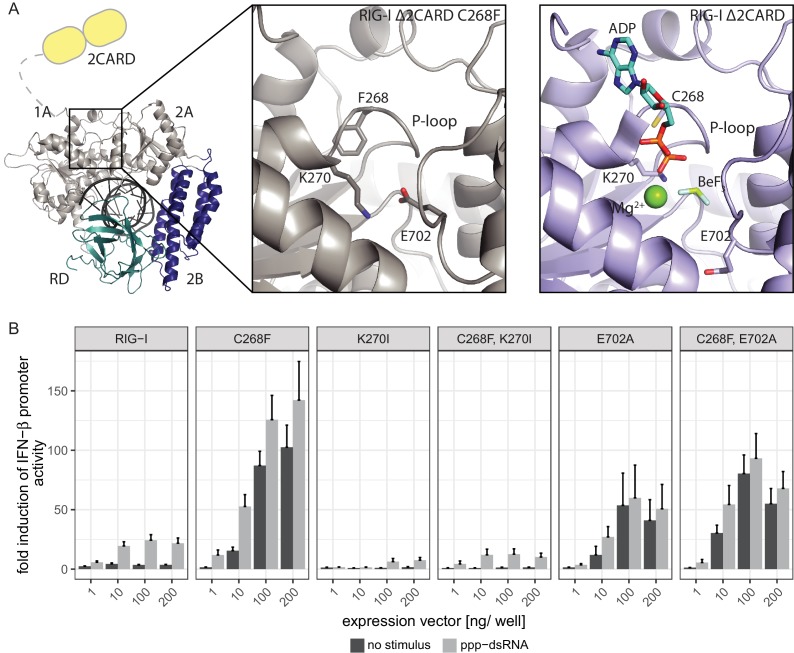
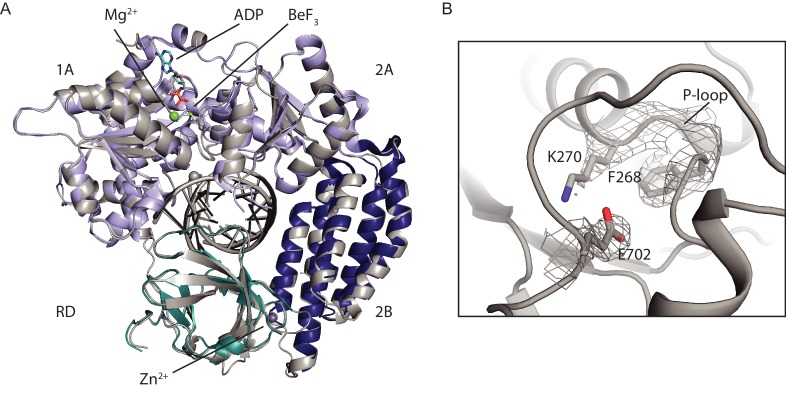
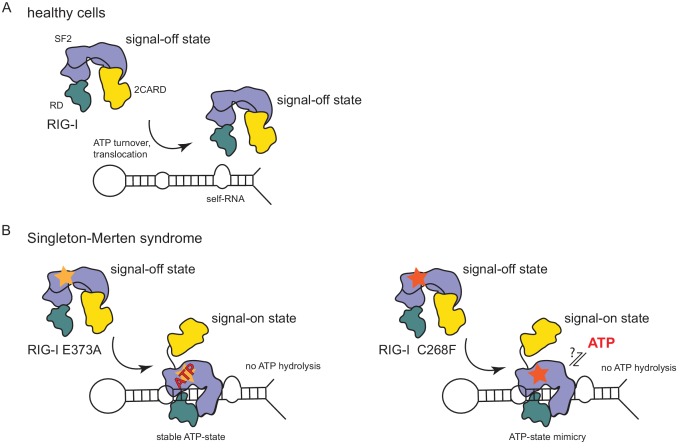
The innate immune sensor retinoic acid-inducible gene I (RIG-I) detects cytosolic viral RNA and requires a conformational change caused by both ATP and RNA binding to induce an active signaling state and to trigger an immune response. Previously, we showed that ATP hydrolysis removes RIG-I from lower-affinity self-RNAs (<xref ref-type="bibr" rid="bib19">Lässig et al., 2015</xref>), revealing how ATP turnover helps RIG-I distinguish viral from self-RNA and explaining why a mutation in a motif that slows down ATP hydrolysis causes the autoimmune disease Singleton-Merten syndrome (SMS). Here we show that a different, mechanistically unexplained SMS variant, C268F, which is localized in the ATP-binding P-loop, can signal independently of ATP but is still dependent on RNA. The structure of RIG-I C268F in complex with double-stranded RNA reveals that C268F helps induce a structural conformation in RIG-I that is similar to that induced by ATP. Our results uncover an unexpected mechanism to explain how a mutation in a P-loop ATPase can induce a gain-of-function ATP state in the absence of ATP.
Keywords: ATPase domain; RIG-I; RLR; Singleton-Merten syndrome; autoimmune response/disease; human; immunology; inflammation; innate immune system; molecular biophysics; structural biology.
© 2018, Lässig et al.
Conflict of interest statement
CL, KL, JG, SM, OF, KH No competing interests declared
Figures
Similar articles
-
Mutations in DDX58, which encodes RIG-I, cause atypical Singleton-Merten syndrome.Am J Hum Genet. 2015 Feb 5;96(2):266-74. doi: 10.1016/j.ajhg.2014.11.019. Epub 2015 Jan 22. Am J Hum Genet. 2015. PMID: 25620203 Free PMC article.
-
A case of Singleton-Merten syndrome without cardiac involvement harboring a novel IFIH1 variant.Am J Med Genet A. 2020 Jun;182(6):1535-1536. doi: 10.1002/ajmg.a.61556. Epub 2020 Mar 23. Am J Med Genet A. 2020. PMID: 32202700 No abstract available.
-
A novel IFIH1 mutation in the pincer domain underlies the clinical features of both Aicardi-Goutières and Singleton-Merten syndromes in a single patient.Br J Dermatol. 2018 Feb;178(2):e111-e113. doi: 10.1111/bjd.15869. Epub 2017 Dec 22. Br J Dermatol. 2018. PMID: 29270977 No abstract available.
-
MDA5-Associated Neuroinflammation and the Singleton-Merten Syndrome: Two Faces of the Same Type I Interferonopathy Spectrum.J Interferon Cytokine Res. 2017 May;37(5):214-219. doi: 10.1089/jir.2017.0004. J Interferon Cytokine Res. 2017. PMID: 28475458 Free PMC article. Review.
-
RNA Editing in Interferonopathies.Methods Mol Biol. 2021;2181:269-286. doi: 10.1007/978-1-0716-0787-9_16. Methods Mol Biol. 2021. PMID: 32729086 Review.
Cited by
-
HDX-MS reveals dysregulated checkpoints that compromise discrimination against self RNA during RIG-I mediated autoimmunity.Nat Commun. 2018 Dec 18;9(1):5366. doi: 10.1038/s41467-018-07780-z. Nat Commun. 2018. PMID: 30560918 Free PMC article.
-
Thymidine phosphorylase facilitates retinoic acid inducible gene-I induced endothelial dysfunction.Cell Death Dis. 2023 Apr 26;14(4):294. doi: 10.1038/s41419-023-05821-0. Cell Death Dis. 2023. PMID: 37100811 Free PMC article.
-
Proofreading mechanisms of the innate immune receptor RIG-I: distinguishing self and viral RNA.Biochem Soc Trans. 2024 Jun 26;52(3):1131-1148. doi: 10.1042/BST20230724. Biochem Soc Trans. 2024. PMID: 38884803 Free PMC article. Review.
-
RNA binding activates RIG-I by releasing an autorepressed signaling domain.Sci Adv. 2019 Oct 2;5(10):eaax3641. doi: 10.1126/sciadv.aax3641. eCollection 2019 Oct. Sci Adv. 2019. PMID: 31616790 Free PMC article.
-
Cellular functions of eukaryotic RNA helicases and their links to human diseases.Nat Rev Mol Cell Biol. 2023 Oct;24(10):749-769. doi: 10.1038/s41580-023-00628-5. Epub 2023 Jul 20. Nat Rev Mol Cell Biol. 2023. PMID: 37474727 Review.
References
-
- Afonine PV, Grosse-Kunstleve RW, Echols N, Headd JJ, Moriarty NW, Mustyakimov M, Terwilliger TC, Urzhumtsev A, Zwart PH, Adams PD. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallographica Section D Biological Crystallography. 2012;68:352–367. doi: 10.1107/S0907444912001308. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- BioSysNet/Bayerisches Staatsministerium für Bildung und Kultus, Wissenschaft und Kunst/International
- CIPSM/Deutsche Forschungsgemeinschaft/International
- HO2489/8/Deutsche Forschungsgemeinschaft/International
- CRC1054 project B02/Deutsche Forschungsgemeinschaft/International
- CRC/TRR 237/Deutsche Forschungsgemeinschaft/International
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
