

Subdiffraction-resolution live-cell imaging for visualizing thylakoid membranes
- PMID: 29982996
- PMCID: PMC6150804
- DOI: 10.1111/tpj.14021
Subdiffraction-resolution live-cell imaging for visualizing thylakoid membranes
Abstract
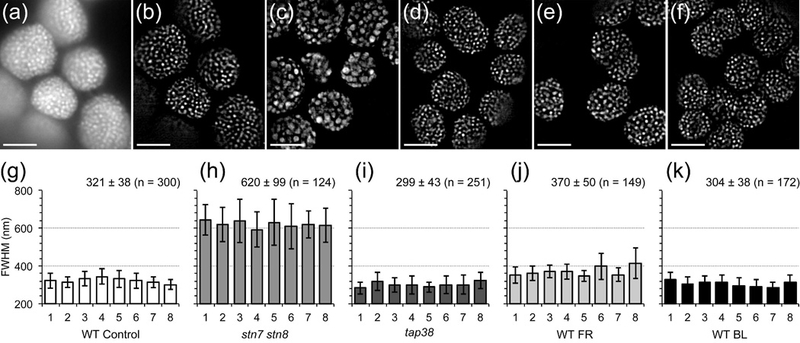
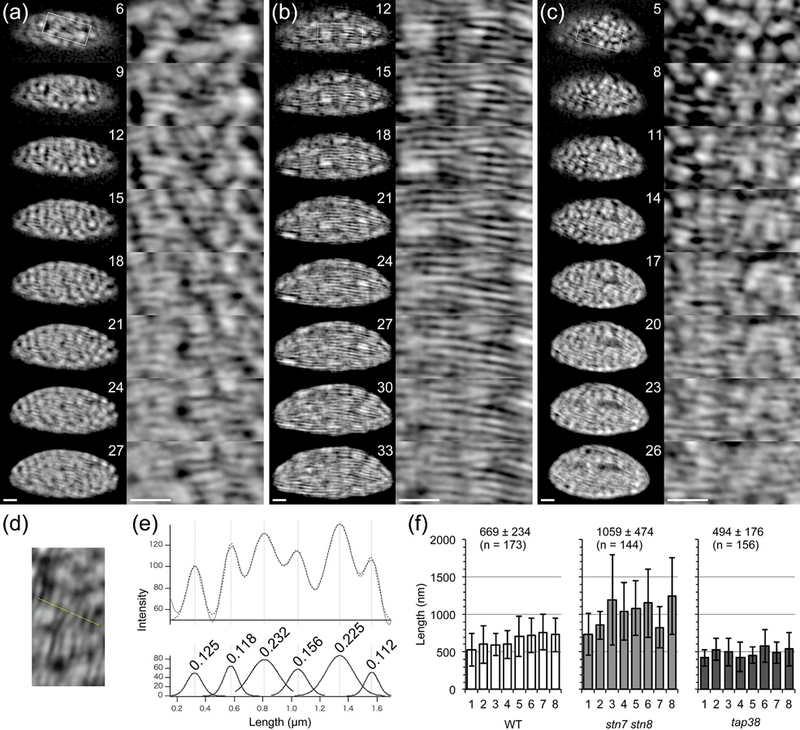
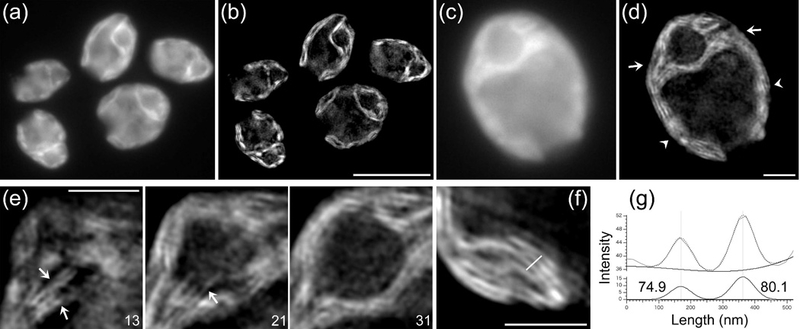
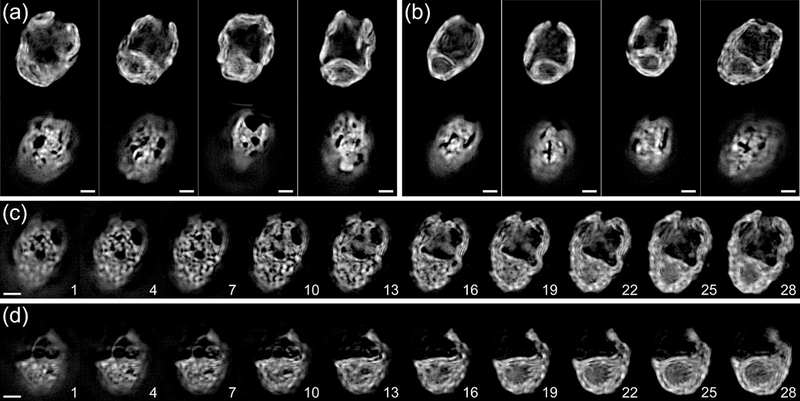
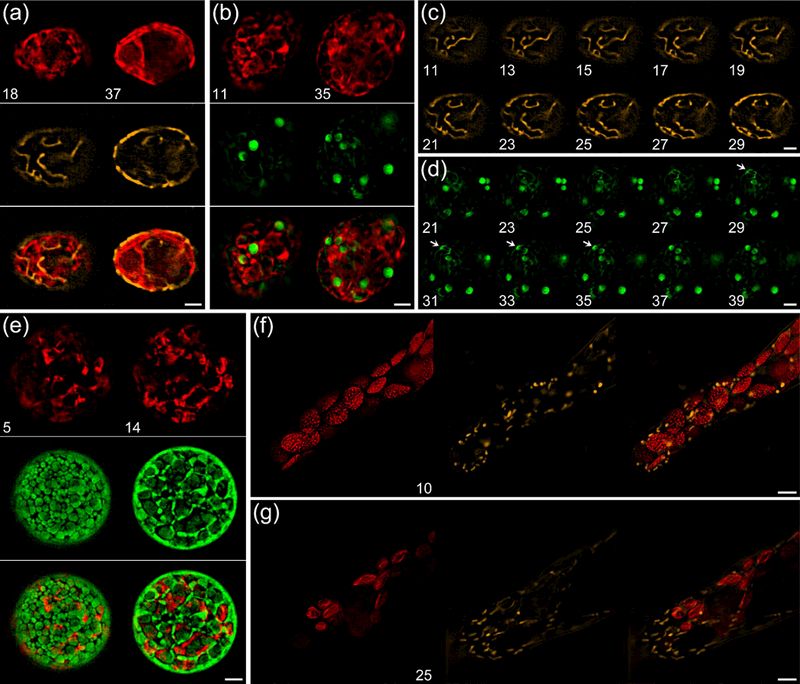
The chloroplast is the chlorophyll-containing organelle that produces energy through photosynthesis. Within the chloroplast is an intricate network of thylakoid membranes containing photosynthetic membrane proteins that mediate electron transport and generate chemical energy. Historically, electron microscopy (EM) has been a powerful tool for visualizing the macromolecular structure and organization of thylakoid membranes. However, an understanding of thylakoid membrane dynamics remains elusive because EM requires fixation and sectioning. To improve our knowledge of thylakoid membrane dynamics we need to consider at least two issues: (i) the live-cell imaging conditions needed to visualize active processes in vivo; and (ii) the spatial resolution required to differentiate the characteristics of thylakoid membranes. Here, we utilize three-dimensional structured illumination microscopy (3D-SIM) to explore the optimal imaging conditions for investigating the dynamics of thylakoid membranes in living plant and algal cells. We show that 3D-SIM is capable of examining broad characteristics of thylakoid structures in chloroplasts of the vascular plant Arabidopsis thaliana and distinguishing the structural differences between wild-type and mutant strains. Using 3D-SIM, we also visualize thylakoid organization in whole cells of the green alga Chlamydomonas reinhardtii. These data reveal that high light intensity changes thylakoid membrane structure in C. reinhardtii. Moreover, we observed the green alga Chromochloris zofingiensis and the moss Physcomitrella patens to show the applicability of 3D-SIM. This study demonstrates that 3D-SIM is a promising approach for studying the dynamics of thylakoid membranes in photoautotrophic organisms during photoacclimation processes.
Keywords: Arabidopsis thaliana; Chlamydomonas reinhardtii; live-cell imaging; structured illumination microscopy; technical advance; thylakoid structure.
© 2018 The Authors The Plant Journal © 2018 John Wiley & Sons Ltd.
Conflict of interest statement
CONFLICT OF INTEREST
The authors declare no conflict of interest.
Figures
Similar articles
-
Visualizing structural dynamics of thylakoid membranes.Sci Rep. 2014 Jan 20;4:3768. doi: 10.1038/srep03768. Sci Rep. 2014. PMID: 24442007 Free PMC article.
-
Charting the native architecture of Chlamydomonas thylakoid membranes with single-molecule precision.Elife. 2020 Apr 16;9:e53740. doi: 10.7554/eLife.53740. Elife. 2020. PMID: 32297859 Free PMC article.
-
Live-cell visualization of excitation energy dynamics in chloroplast thylakoid structures.Sci Rep. 2016 Jul 15;6:29940. doi: 10.1038/srep29940. Sci Rep. 2016. PMID: 27416900 Free PMC article.
-
Electron tomography of plant thylakoid membranes.J Exp Bot. 2011 Apr;62(7):2393-402. doi: 10.1093/jxb/err034. Epub 2011 Mar 25. J Exp Bot. 2011. PMID: 21441405 Review.
-
Biogenesis and origin of thylakoid membranes.Biochim Biophys Acta. 2001 Dec 12;1541(1-2):91-101. doi: 10.1016/s0167-4889(01)00153-7. Biochim Biophys Acta. 2001. PMID: 11750665 Review.
Cited by
-
Single chloroplast in folio imaging sheds light on photosystem energy redistribution during state transitions.Plant Physiol. 2023 Feb 12;191(2):1186-1198. doi: 10.1093/plphys/kiac561. Plant Physiol. 2023. PMID: 36478277 Free PMC article.
-
Action of 2,6-Dichloro-1,4-benzoquinone on the O2-Evolving Activity of Photosystem II in Chlamydomonas reinhardtii Cells with and without Cell Wall: Inhibitory Effect of Its Oxidized Form.Cells. 2023 Mar 15;12(6):907. doi: 10.3390/cells12060907. Cells. 2023. PMID: 36980248 Free PMC article.
-
The Role of Phosphorylation Dynamics of CURVATURE THYLAKOID 1B in Plant Thylakoid Membranes.Plant Physiol. 2019 Dec;181(4):1615-1631. doi: 10.1104/pp.19.00942. Epub 2019 Oct 15. Plant Physiol. 2019. PMID: 31615849 Free PMC article.
-
How to Measure Grana - Ultrastructural Features of Thylakoid Membranes of Plant Chloroplasts.Front Plant Sci. 2021 Oct 6;12:756009. doi: 10.3389/fpls.2021.756009. eCollection 2021. Front Plant Sci. 2021. PMID: 34691132 Free PMC article. Review.
-
Atomic Force Microscopy Visualizes Mobility of Photosynthetic Proteins in Grana Thylakoid Membranes.Biophys J. 2020 Apr 21;118(8):1876-1886. doi: 10.1016/j.bpj.2020.02.029. Epub 2020 Mar 13. Biophys J. 2020. PMID: 32224302 Free PMC article.
References
-
- Armbruster U , Labs M , Pribil M , Viola S , Xu W , Scharfenberg M , Hertle AP , Rojahn U , Jensen PE , Rappaport F , Joliot P , Dormann P , Wanner G and Leister D (2013) Arabidopsis CURVATURE THYLAKOID1 proteins modify thylakoid architecture by inducing membrane curvature. Plant Cell, 25, 2661–2678. - PMC - PubMed
-
- Ballottari M , Truong TB , De Re E , Erickson E , Stella GR , Fleming GR , Bassi R and Niyogi KK (2016) Identification of pH-sensing sites in the light harvesting complex stress-related 3 protein essential for triggering non-photochemical quenching in Chlamydomonas reinhardtii. J. Biol. Chem, 291, 7334–7346. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
