

Rab18 is not necessary for lipid droplet biogenesis or turnover in human mammary carcinoma cells
- PMID: 29949452
- PMCID: PMC6232964
- DOI: 10.1091/mbc.E18-05-0282
Rab18 is not necessary for lipid droplet biogenesis or turnover in human mammary carcinoma cells
Abstract
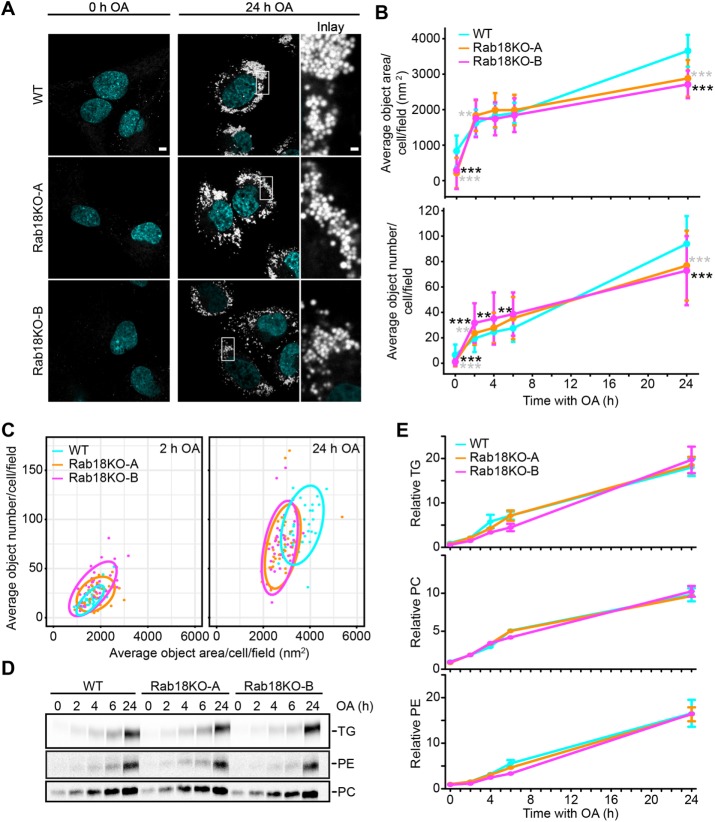
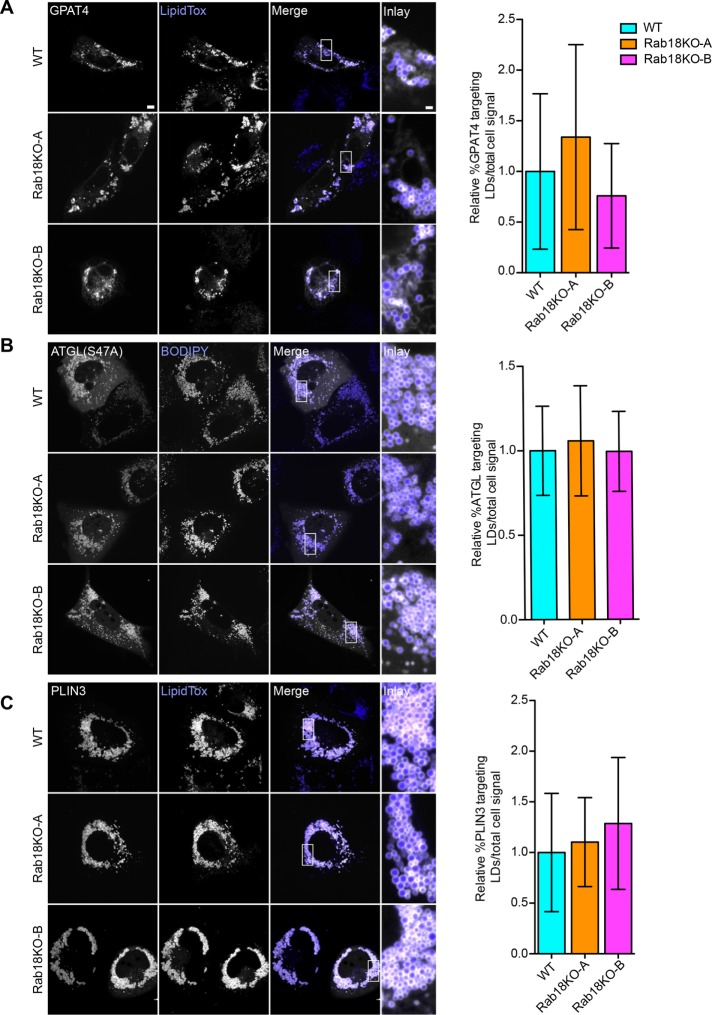
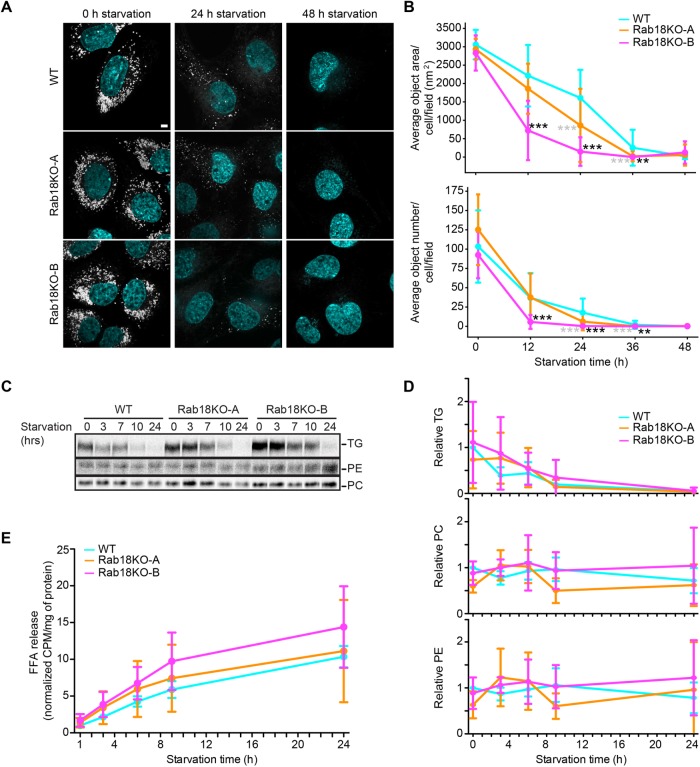
Rab GTPases recruit peripheral membrane proteins and can define organelle identity. Rab18 localizes to the endoplasmic reticulum (ER) but also to lipid droplets (LDs), where it has been implicated in effector protein recruitment and in defining LD identity. Here, we studied Rab18 localization and function in a human mammary carcinoma cell line. Rab18 localized to the ER and to LD membranes on LD induction, with the latter depending on the Rab18 activation state. In cells lacking Rab18, LDs were modestly reduced in size and numbers, but we found little evidence for Rab18 function in LD formation, LD turnover on cell starvation, or the targeting of several proteins to LDs. We conclude that Rab18 is not a general, necessary component of the protein machinery involved in LD biogenesis or turnover.
Figures


Similar articles
-
Rab18 promotes lipid droplet (LD) growth by tethering the ER to LDs through SNARE and NRZ interactions.J Cell Biol. 2018 Mar 5;217(3):975-995. doi: 10.1083/jcb.201704184. Epub 2018 Jan 24. J Cell Biol. 2018. PMID: 29367353 Free PMC article.
-
Rab18 binds PLIN2 and ACSL3 to mediate lipid droplet dynamics.Biochim Biophys Acta Mol Cell Biol Lipids. 2021 Jul;1866(7):158923. doi: 10.1016/j.bbalip.2021.158923. Epub 2021 Mar 11. Biochim Biophys Acta Mol Cell Biol Lipids. 2021. PMID: 33713834
-
Rab18 localizes to lipid droplets and induces their close apposition to the endoplasmic reticulum-derived membrane.J Cell Sci. 2005 Jun 15;118(Pt 12):2601-11. doi: 10.1242/jcs.02401. Epub 2005 May 24. J Cell Sci. 2005. PMID: 15914536
-
Come a little bit closer! Lipid droplet-ER contact sites are getting crowded.Biochim Biophys Acta Mol Cell Res. 2020 Feb;1867(2):118603. doi: 10.1016/j.bbamcr.2019.118603. Epub 2019 Nov 13. Biochim Biophys Acta Mol Cell Res. 2020. PMID: 31733263 Review.
-
Interactions of Lipid Droplets with the Intracellular Transport Machinery.Int J Mol Sci. 2021 Mar 9;22(5):2776. doi: 10.3390/ijms22052776. Int J Mol Sci. 2021. PMID: 33803444 Free PMC article. Review.
Cited by
-
Porcine reproductive and respiratory syndrome virus 2 hijacks CMA-mediated lipolysis through upregulation of small GTPase RAB18.PLoS Pathog. 2024 Apr 12;20(4):e1012123. doi: 10.1371/journal.ppat.1012123. eCollection 2024 Apr. PLoS Pathog. 2024. PMID: 38607975 Free PMC article.
-
Dietary S. maltophilia induces supersized lipid droplets by enhancing lipogenesis and ER-LD contacts in C. elegans.Gut Microbes. 2022 Jan-Dec;14(1):2013762. doi: 10.1080/19490976.2021.2013762. Gut Microbes. 2022. PMID: 35112996 Free PMC article.
-
Identification of novel lipid droplet factors that regulate lipophagy and cholesterol efflux in macrophage foam cells.Autophagy. 2021 Nov;17(11):3671-3689. doi: 10.1080/15548627.2021.1886839. Epub 2021 Feb 26. Autophagy. 2021. PMID: 33590792 Free PMC article.
-
A Unique Junctional Interface at Contact Sites Between the Endoplasmic Reticulum and Lipid Droplets.Front Cell Dev Biol. 2021 Apr 8;9:650186. doi: 10.3389/fcell.2021.650186. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33898445 Free PMC article. Review.
-
Rab18 Drift in Lipid Droplet and Endoplasmic Reticulum Interactions of Adipocytes under Obesogenic Conditions.Int J Mol Sci. 2023 Dec 6;24(24):17177. doi: 10.3390/ijms242417177. Int J Mol Sci. 2023. PMID: 38139006 Free PMC article.
References
-
- Carpanini SM, McKie L, Thomson D, Wright AK, Gordon SL, Roche SL, Handley MT, Morrison H, Brownstein D, Wishart TM, et al. (2014). A novel mouse model of Warburg Micro syndrome reveals roles for RAB18 in eye development and organisation of the neuronal cytoskeleton. Dis Model Mech , 711–722. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
