

Adjustable viscoelasticity allows for efficient collective cell migration
- PMID: 29859995
- PMCID: PMC6854469
- DOI: 10.1016/j.semcdb.2018.05.027
Adjustable viscoelasticity allows for efficient collective cell migration
Abstract
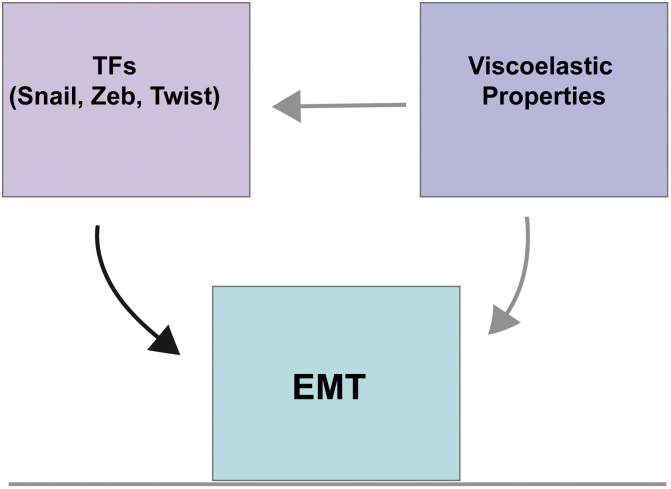
Cell migration is essential for a wide range of biological processes such as embryo morphogenesis, wound healing, regeneration, and also in pathological conditions, such as cancer. In such contexts, cells are required to migrate as individual entities or as highly coordinated collectives, both of which requiring cells to respond to molecular and mechanical cues from their environment. However, whilst the function of chemical cues in cell migration is comparatively well understood, the role of tissue mechanics on cell migration is just starting to be studied. Recent studies suggest that the dynamic tuning of the viscoelasticity within a migratory cluster of cells, and the adequate elastic properties of its surrounding tissues, are essential to allow efficient collective cell migration in vivo. In this review we focus on the role of viscoelasticity in the control of collective cell migration in various cellular systems, mentioning briefly some aspects of single cell migration. We aim to provide details on how viscoelasticity of collectively migrating groups of cells and their surroundings is adjusted to ensure correct morphogenesis, wound healing, and metastasis. Finally, we attempt to show that environmental viscoelasticity triggers molecular changes within migrating clusters and that these new molecular setups modify clusters' viscoelasticity, ultimately allowing them to migrate across the challenging geometries of their microenvironment.
Keywords: Adherens junctions; Cancer invasion; Collective migration; EMT; Mechanical microenvironment; Viscoelasticity.
Copyright © 2018 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
Similar articles
-
Long-time viscoelasticity of multicellular surfaces caused by collective cell migration - Multi-scale modeling considerations.Semin Cell Dev Biol. 2019 Sep;93:87-96. doi: 10.1016/j.semcdb.2018.08.002. Epub 2018 Aug 7. Semin Cell Dev Biol. 2019. PMID: 30086376 Review.
-
Viscoelasticity of multicellular systems caused by collective cell migration: dynamics at the biointerface.Eur Biophys J. 2020 May;49(3-4):253-265. doi: 10.1007/s00249-020-01431-2. Epub 2020 May 10. Eur Biophys J. 2020. PMID: 32390090
-
Collective cell migration and residual stress accumulation: Rheological consideration.J Biomech. 2020 Jul 17;108:109898. doi: 10.1016/j.jbiomech.2020.109898. Epub 2020 Jun 13. J Biomech. 2020. PMID: 32636009 Review.
-
Tissue stiffening coordinates morphogenesis by triggering collective cell migration in vivo.Nature. 2018 Feb 22;554(7693):523-527. doi: 10.1038/nature25742. Epub 2018 Feb 14. Nature. 2018. PMID: 29443958 Free PMC article.
-
Collective cell migration: Implications for wound healing and cancer invasion.Burns Trauma. 2013 Jun 18;1(1):21-6. doi: 10.4103/2321-3868.113331. eCollection 2013. Burns Trauma. 2013. PMID: 27574618 Free PMC article.
Cited by
-
Mechanical regulation of the early stages of angiogenesis.J R Soc Interface. 2022 Dec;19(197):20220360. doi: 10.1098/rsif.2022.0360. Epub 2022 Dec 7. J R Soc Interface. 2022. PMID: 36475392 Free PMC article. Review.
-
The extracellular matrix in development.Development. 2020 May 28;147(10):dev175596. doi: 10.1242/dev.175596. Development. 2020. PMID: 32467294 Free PMC article. Review.
-
Mechanosensory feedback loops during chronic inflammation.Front Cell Dev Biol. 2023 Jul 10;11:1225677. doi: 10.3389/fcell.2023.1225677. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37492225 Free PMC article. Review.
-
Viscoelastic Biomaterials for Tissue Regeneration.Tissue Eng Part C Methods. 2022 Jul;28(7):289-300. doi: 10.1089/ten.TEC.2022.0040. Tissue Eng Part C Methods. 2022. PMID: 35442107 Free PMC article. Review.
-
Tubulin acetylation promotes penetrative capacity of cells undergoing radial intercalation.Cell Rep. 2021 Aug 17;36(7):109556. doi: 10.1016/j.celrep.2021.109556. Cell Rep. 2021. PMID: 34407402 Free PMC article.
References
-
- Mayor R., Etienne-Manneville S. The front and rear of collective cell migration. Nat. Rev. Mol. Cell Biol. 2016;17(2):97–109. - PubMed
-
- Friedl P., Gilmour D. Collective cell migration in morphogenesis, regeneration and cancer. Nat. Rev. Mol. Cell. Biol. 2009;10(7):445–457. - PubMed
-
- Rorth P. Collective cell migration. Annu Rev. Cell Dev. Biol. 2009;25:407–429. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
