

Early Seizures Prematurely Unsilence Auditory Synapses to Disrupt Thalamocortical Critical Period Plasticity
- PMID: 29847785
- PMCID: PMC6446922
- DOI: 10.1016/j.celrep.2018.04.108
Early Seizures Prematurely Unsilence Auditory Synapses to Disrupt Thalamocortical Critical Period Plasticity
Abstract
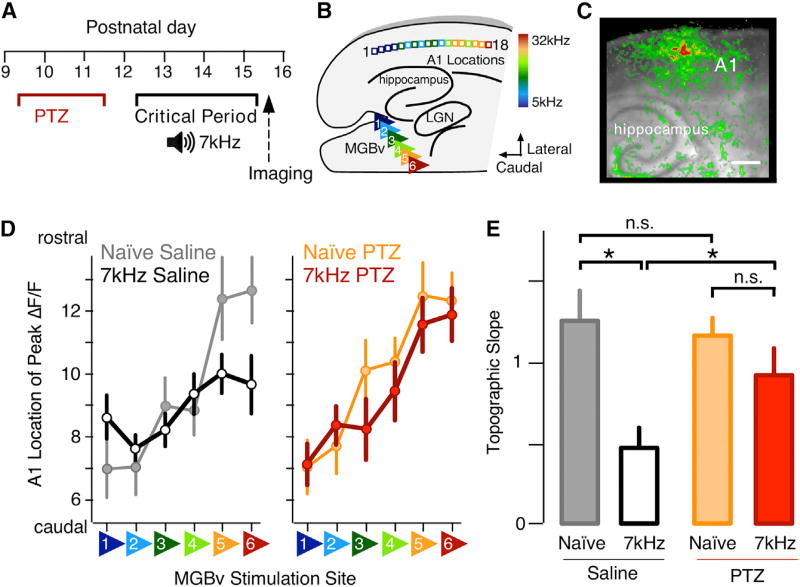
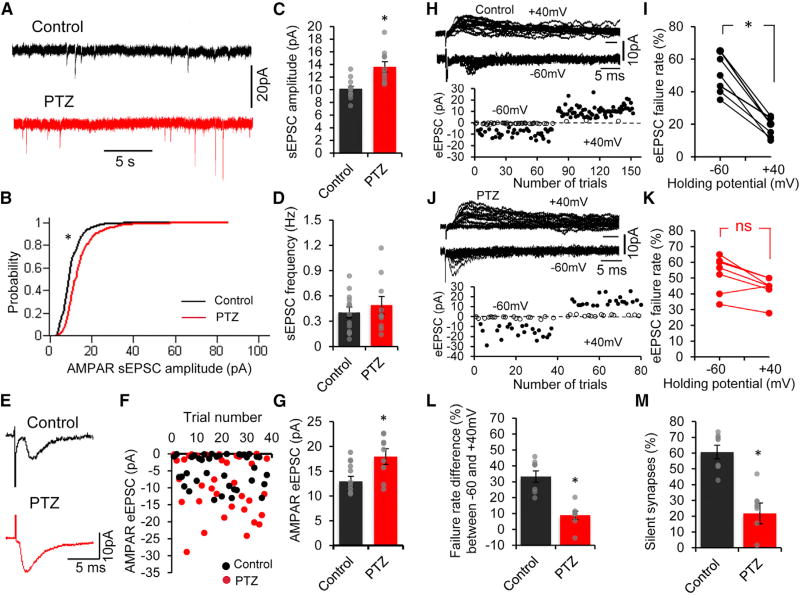
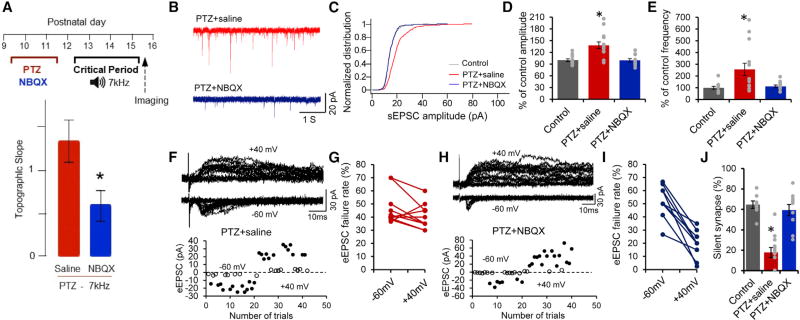
Heightened neural excitability in infancy and childhood results in increased susceptibility to seizures. Such early-life seizures are associated with language deficits and autism that can result from aberrant development of the auditory cortex. Here, we show that early-life seizures disrupt a critical period (CP) for tonotopic map plasticity in primary auditory cortex (A1). We show that this CP is characterized by a prevalence of "silent," NMDA-receptor (NMDAR)-only, glutamate receptor synapses in auditory cortex that become "unsilenced" due to activity-dependent AMPA receptor (AMPAR) insertion. Induction of seizures prior to this CP occludes tonotopic map plasticity by prematurely unsilencing NMDAR-only synapses. Further, brief treatment with the AMPAR antagonist NBQX following seizures, prior to the CP, prevents synapse unsilencing and permits subsequent A1 plasticity. These findings reveal that early-life seizures modify CP regulators and suggest that therapeutic targets for early post-seizure treatment can rescue CP plasticity.
Keywords: AMPA receptor; NBQX; NMDA receptor; auditory cortex; autism; development; epilepsy; neurodevelopmental disorders; silent synapses; tonotopic plasticity.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

Comment in
-
"Can You Hear Me Now?" AMPA Receptor-Mediated Tonotopy Disruption by Early Life Seizures.Epilepsy Curr. 2018 Nov-Dec;18(6):391-393. doi: 10.5698/1535-7597.18.6.391. Epilepsy Curr. 2018. PMID: 30568558 Free PMC article. No abstract available.
Similar articles
-
Early-life seizures alter synaptic calcium-permeable AMPA receptor function and plasticity.Mol Cell Neurosci. 2016 Oct;76:11-20. doi: 10.1016/j.mcn.2016.08.002. Epub 2016 Aug 10. Mol Cell Neurosci. 2016. PMID: 27521497 Free PMC article.
-
AMPA receptor antagonist NBQX attenuates later-life epileptic seizures and autistic-like social deficits following neonatal seizures.Epilepsia. 2013 Nov;54(11):1922-32. doi: 10.1111/epi.12378. Epub 2013 Oct 1. Epilepsia. 2013. PMID: 24117347 Free PMC article.
-
Barrel cortex critical period plasticity is independent of changes in NMDA receptor subunit composition.Neuron. 2001 Nov 20;32(4):619-34. doi: 10.1016/s0896-6273(01)00501-3. Neuron. 2001. PMID: 11719203
-
Synaptic plasticity at thalamocortical synapses in developing rat somatosensory cortex: LTP, LTD, and silent synapses.J Neurobiol. 1999 Oct;41(1):92-101. J Neurobiol. 1999. PMID: 10504196 Review.
-
Silent synapse: A new player in visual cortex critical period plasticity.Pharmacol Res. 2019 Mar;141:586-590. doi: 10.1016/j.phrs.2019.01.031. Epub 2019 Jan 16. Pharmacol Res. 2019. PMID: 30659896 Review.
Cited by
-
Early-life adversity and neurological disease: age-old questions and novel answers.Nat Rev Neurol. 2019 Nov;15(11):657-669. doi: 10.1038/s41582-019-0246-5. Epub 2019 Sep 17. Nat Rev Neurol. 2019. PMID: 31530940 Free PMC article. Review.
-
Enduring memory consequences of early-life stress / adversity: Structural, synaptic, molecular and epigenetic mechanisms.Neurobiol Stress. 2024 Aug 30;33:100669. doi: 10.1016/j.ynstr.2024.100669. eCollection 2024 Nov. Neurobiol Stress. 2024. PMID: 39309367 Free PMC article. Review.
-
The influence of unpredictable, fragmented parental signals on the developing brain.Front Neuroendocrinol. 2019 Apr;53:100736. doi: 10.1016/j.yfrne.2019.01.002. Epub 2019 Jan 31. Front Neuroendocrinol. 2019. PMID: 30711600 Free PMC article. Review.
-
Occlusion of activity dependent synaptic plasticity by late hypoxic long term potentiation after neonatal intermittent hypoxia.Exp Neurol. 2021 Mar;337:113575. doi: 10.1016/j.expneurol.2020.113575. Epub 2020 Dec 21. Exp Neurol. 2021. PMID: 33358869 Free PMC article. Review.
-
Dysregulation of GABAA Receptor-Mediated Neurotransmission during the Auditory Cortex Critical Period in the Fragile X Syndrome Mouse Model.Cereb Cortex. 2021 Nov 23;32(1):197-215. doi: 10.1093/cercor/bhab203. Cereb Cortex. 2021. PMID: 34223875 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
