

PUF60-activated exons uncover altered 3' splice-site selection by germline missense mutations in a single RRM
- PMID: 29788428
- PMCID: PMC6093180
- DOI: 10.1093/nar/gky389
PUF60-activated exons uncover altered 3' splice-site selection by germline missense mutations in a single RRM
Abstract
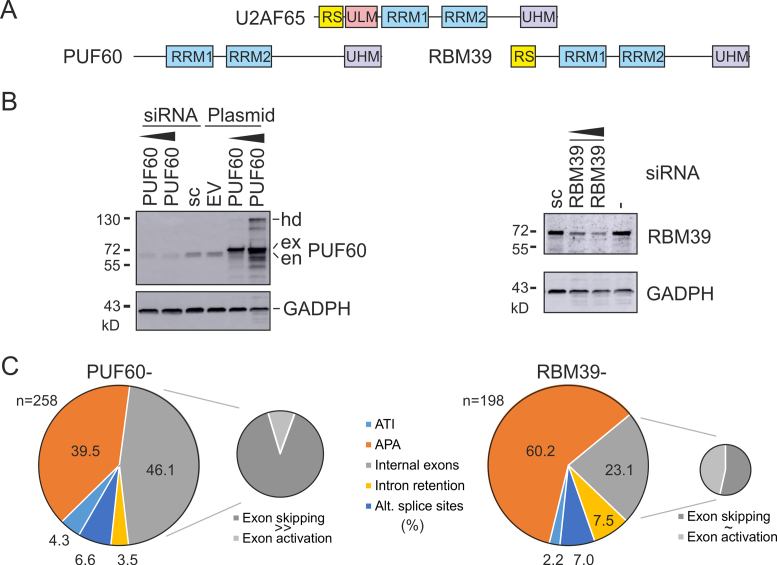
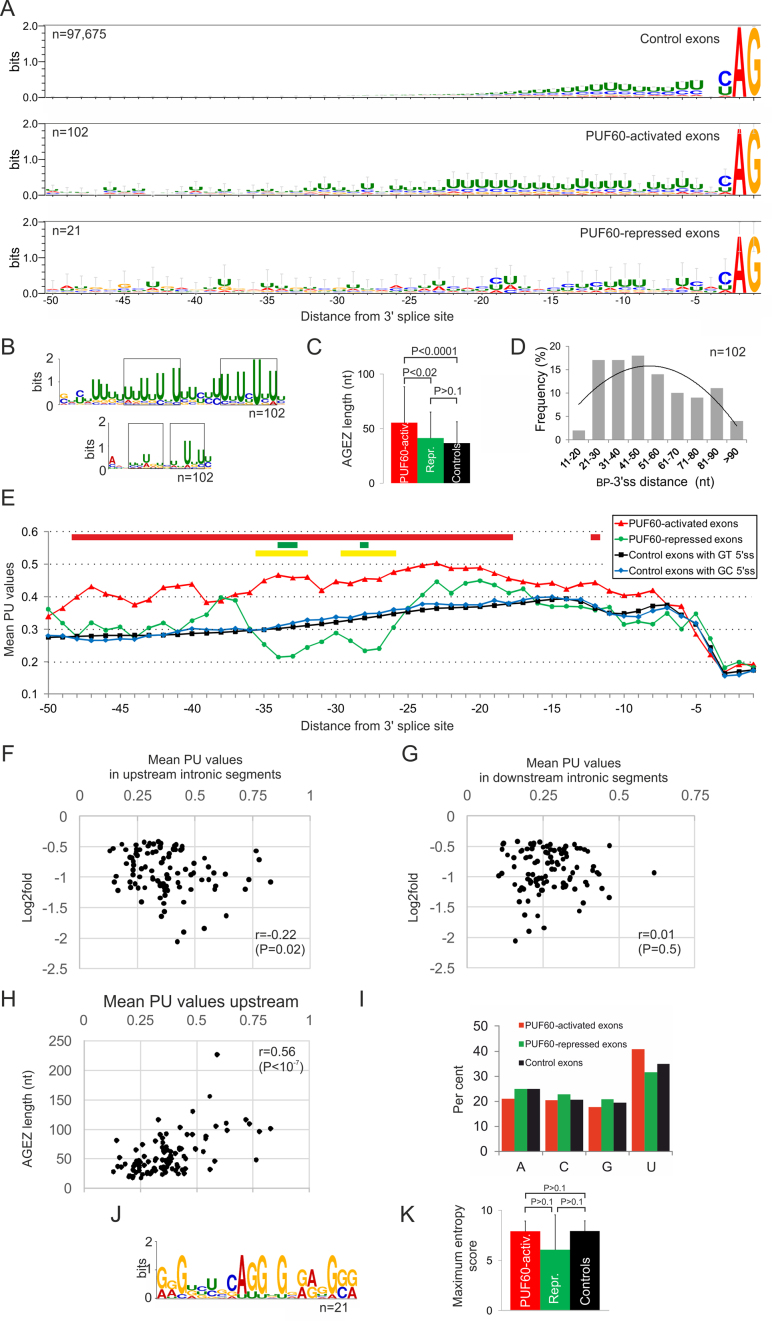
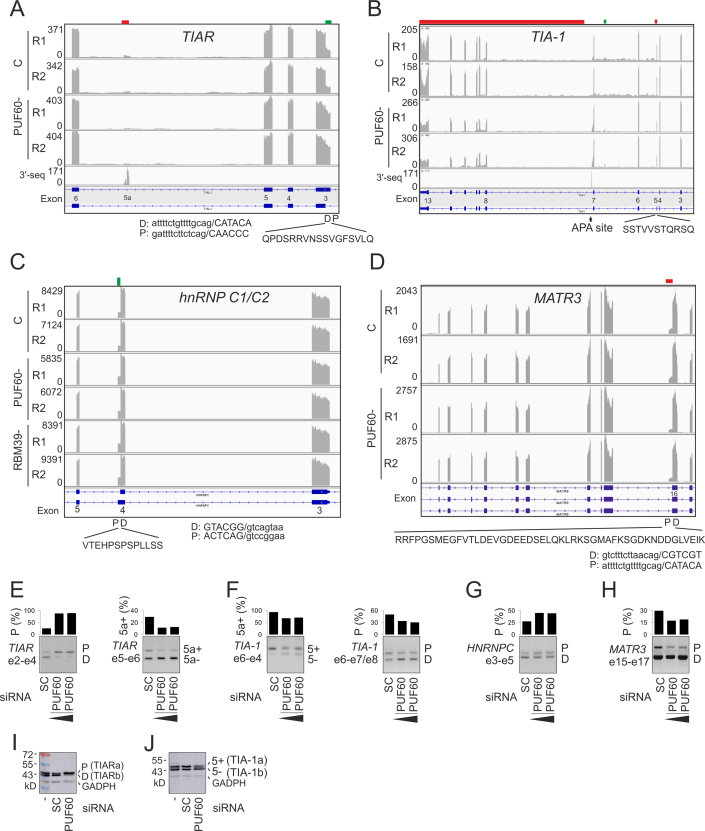
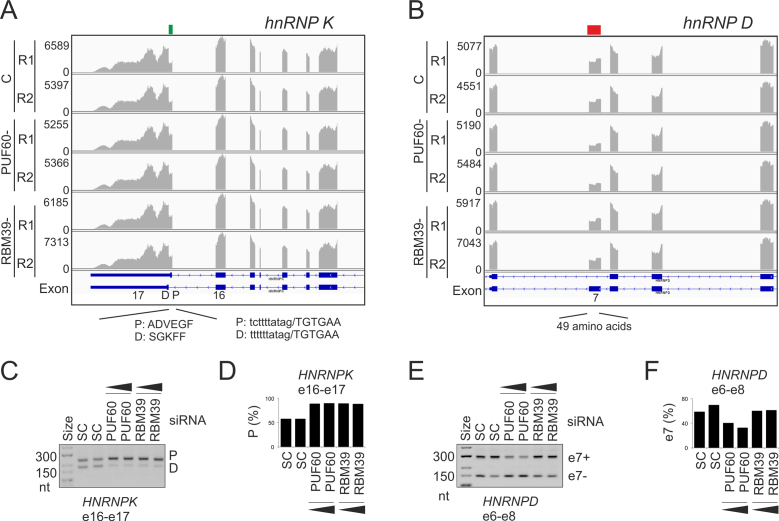
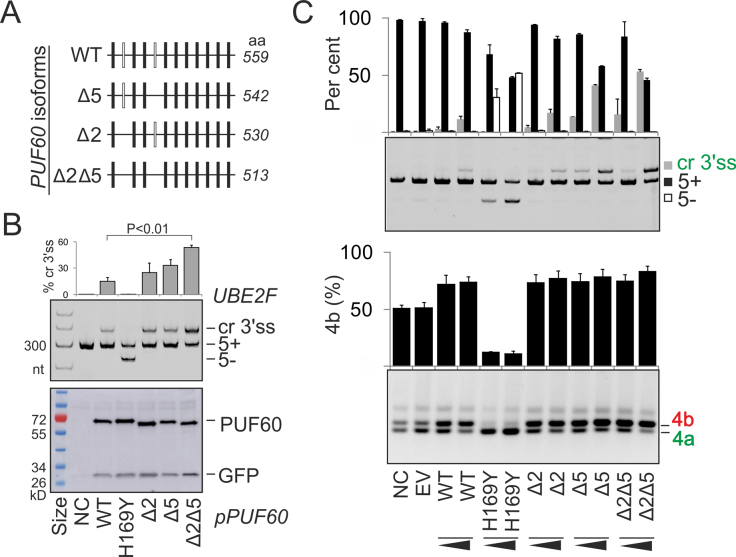
PUF60 is a splicing factor that binds uridine (U)-rich tracts and facilitates association of the U2 small nuclear ribonucleoprotein with primary transcripts. PUF60 deficiency (PD) causes a developmental delay coupled with intellectual disability and spinal, cardiac, ocular and renal defects, but PD pathogenesis is not understood. Using RNA-Seq, we identify human PUF60-regulated exons and show that PUF60 preferentially acts as their activator. PUF60-activated internal exons are enriched for Us upstream of their 3' splice sites (3'ss), are preceded by longer AG dinucleotide exclusion zones and more distant branch sites, with a higher probability of unpaired interactions across a typical branch site location as compared to control exons. In contrast, PUF60-repressed exons show U-depletion with lower estimates of RNA single-strandedness. We also describe PUF60-regulated, alternatively spliced isoforms encoding other U-bound splicing factors, including PUF60 partners, suggesting that they are co-regulated in the cell, and identify PUF60-regulated exons derived from transposed elements. PD-associated amino-acid substitutions, even within a single RNA recognition motif (RRM), altered selection of competing 3'ss and branch points of a PUF60-dependent exon and the 3'ss choice was also influenced by alternative splicing of PUF60. Finally, we propose that differential distribution of RNA processing steps detected in cells lacking PUF60 and the PUF60-paralog RBM39 is due to the RBM39 RS domain interactions. Together, these results provide new insights into regulation of exon usage by the 3'ss organization and reveal that germline mutation heterogeneity in RRMs can enhance phenotypic variability at the level of splice-site and branch-site selection.
Figures
Similar articles
-
Cancer-Associated Substitutions in RNA Recognition Motifs of PUF60 and U2AF65 Reveal Residues Required for Correct Folding and 3' Splice-Site Selection.Cancers (Basel). 2020 Jul 11;12(7):1865. doi: 10.3390/cancers12071865. Cancers (Basel). 2020. PMID: 32664474 Free PMC article.
-
Identification of U2AF(35)-dependent exons by RNA-Seq reveals a link between 3' splice-site organization and activity of U2AF-related proteins.Nucleic Acids Res. 2015 Apr 20;43(7):3747-63. doi: 10.1093/nar/gkv194. Epub 2015 Mar 16. Nucleic Acids Res. 2015. PMID: 25779042 Free PMC article.
-
Splicing Enhancers at Intron-Exon Borders Participate in Acceptor Splice Sites Recognition.Int J Mol Sci. 2020 Sep 8;21(18):6553. doi: 10.3390/ijms21186553. Int J Mol Sci. 2020. PMID: 32911621 Free PMC article.
-
Diverse regulation of 3' splice site usage.Cell Mol Life Sci. 2015 Dec;72(24):4771-93. doi: 10.1007/s00018-015-2037-5. Epub 2015 Sep 14. Cell Mol Life Sci. 2015. PMID: 26370726 Free PMC article. Review.
-
Splicing at the phase-separated nuclear speckle interface: a model.Nucleic Acids Res. 2021 Jan 25;49(2):636-645. doi: 10.1093/nar/gkaa1209. Nucleic Acids Res. 2021. PMID: 33337476 Free PMC article. Review.
Cited by
-
Arginine reprograms metabolism in liver cancer via RBM39.Cell. 2023 Nov 9;186(23):5068-5083.e23. doi: 10.1016/j.cell.2023.09.011. Epub 2023 Oct 6. Cell. 2023. PMID: 37804830 Free PMC article.
-
Alu RNA fold links splicing with signal recognition particle proteins.Nucleic Acids Res. 2023 Aug 25;51(15):8199-8216. doi: 10.1093/nar/gkad500. Nucleic Acids Res. 2023. PMID: 37309897 Free PMC article.
-
RNA-binding protein 39: a promising therapeutic target for cancer.Cell Death Discov. 2021 Aug 13;7(1):214. doi: 10.1038/s41420-021-00598-7. Cell Death Discov. 2021. PMID: 34389703 Free PMC article. Review.
-
Decreased spliceosome fidelity and egl-8 intron retention inhibit mTORC1 signaling to promote longevity.Nat Aging. 2022 Sep;2(9):796-808. doi: 10.1038/s43587-022-00275-z. Epub 2022 Sep 19. Nat Aging. 2022. PMID: 37118503 Free PMC article.
-
Identification of spliceosome components pivotal to breast cancer survival.RNA Biol. 2021 Jun;18(6):833-842. doi: 10.1080/15476286.2020.1822636. Epub 2020 Sep 23. RNA Biol. 2021. PMID: 32965163 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
