

Regulation of macrophage immunometabolism in atherosclerosis
- PMID: 29777212
- PMCID: PMC6314674
- DOI: 10.1038/s41590-018-0113-3
Regulation of macrophage immunometabolism in atherosclerosis
Abstract
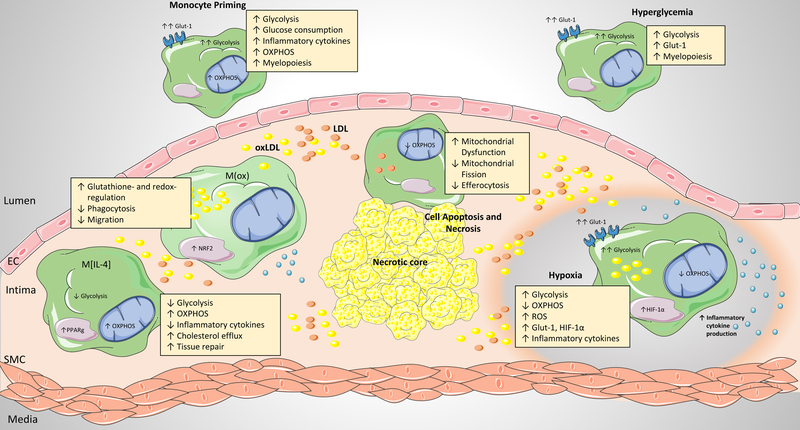
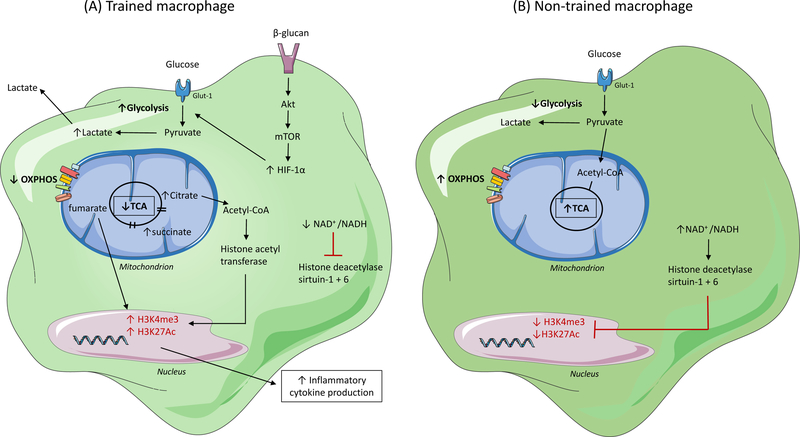
After activation, cells of the myeloid lineage undergo robust metabolic transitions, as well as discrete epigenetic changes, that can dictate both ongoing and future inflammatory responses. In atherosclerosis, in which macrophages play central roles in the initiation, growth, and ultimately rupture of arterial plaques, altered metabolism is a key feature that dictates macrophage function and subsequent disease progression. This Review explores how factors central to the plaque microenvironment (for example, altered cholesterol metabolism, oxidative stress, hypoxia, apoptotic and necrotic cells, and hyperglycemia) shape the metabolic rewiring of macrophages in atherosclerosis as well as how these metabolic shifts in turn alter macrophage immune-effector and tissue-reparative functions. Finally, this overview offers insight into the challenges and opportunities of harnessing metabolism to modulate aberrant macrophage responses in disease.
Figures

Similar articles
-
Targeting Macrophage Phenotypes and Metabolism as Novel Therapeutic Approaches in Atherosclerosis and Related Cardiovascular Diseases.Curr Atheroscler Rep. 2024 Oct;26(10):573-588. doi: 10.1007/s11883-024-01229-z. Epub 2024 Aug 12. Curr Atheroscler Rep. 2024. PMID: 39133247 Free PMC article. Review.
-
Intracellular and Intercellular Aspects of Macrophage Immunometabolism in Atherosclerosis.Circ Res. 2020 Apr 24;126(9):1209-1227. doi: 10.1161/CIRCRESAHA.119.315939. Epub 2020 Apr 23. Circ Res. 2020. PMID: 32324504 Free PMC article. Review.
-
Monocyte and macrophage immunometabolism in atherosclerosis.Semin Immunopathol. 2018 Feb;40(2):203-214. doi: 10.1007/s00281-017-0656-7. Epub 2017 Oct 2. Semin Immunopathol. 2018. PMID: 28971272 Free PMC article. Review.
-
Mitochondria orchestrate macrophage effector functions in atherosclerosis.Mol Aspects Med. 2021 Feb;77:100922. doi: 10.1016/j.mam.2020.100922. Epub 2020 Nov 6. Mol Aspects Med. 2021. PMID: 33162108 Review.
-
Immunometabolism orchestrates training of innate immunity in atherosclerosis.Cardiovasc Res. 2019 Jul 1;115(9):1416-1424. doi: 10.1093/cvr/cvz107. Cardiovasc Res. 2019. PMID: 31050710 Free PMC article. Review.
Cited by
-
Butyrate suppresses atherosclerotic inflammation by regulating macrophages and polarization via GPR43/HDAC-miRNAs axis in ApoE-/- mice.PLoS One. 2023 Mar 8;18(3):e0282685. doi: 10.1371/journal.pone.0282685. eCollection 2023. PLoS One. 2023. PMID: 36888629 Free PMC article.
-
Heme-heme oxygenase-2 reduces the atherosclerosis by preventing inflammation.Curr Res Pharmacol Drug Discov. 2022 Dec 12;4:100141. doi: 10.1016/j.crphar.2022.100141. eCollection 2023. Curr Res Pharmacol Drug Discov. 2022. PMID: 36593926 Free PMC article.
-
A Systematic Review on the Research Progress on Polysaccharides from Fungal Traditional Chinese Medicine.Molecules. 2023 Sep 27;28(19):6816. doi: 10.3390/molecules28196816. Molecules. 2023. PMID: 37836659 Free PMC article. Review.
-
The myocardial infarction-associated transcript 2 inhibits lipid accumulation and promotes cholesterol efflux in oxidized low-density lipoprotein-induced THP-1-derived macrophages via inhibiting mitogen-activated protein kinase signaling and activating the nuclear factor erythroid-related factor 2 signaling pathway.Bioengineered. 2022 Jan;13(1):407-417. doi: 10.1080/21655979.2021.2005932. Bioengineered. 2022. Retraction in: Bioengineered. 2024 Dec;15(1):2299617. doi: 10.1080/21655979.2024.2299617. Retraction in: Bioengineered. 2024 Dec;15(1):2299530. doi: 10.1080/21655979.2024.2299530. PMID: 34783297 Free PMC article. Retracted.
-
Proteomics based identification of KDM5 histone demethylases associated with cardiovascular disease.EBioMedicine. 2019 Mar;41:91-104. doi: 10.1016/j.ebiom.2019.02.040. Epub 2019 Feb 28. EBioMedicine. 2019. PMID: 30826357 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
