

Transmembrane Protein pUL50 of Human Cytomegalovirus Inhibits ISGylation by Downregulating UBE1L
- PMID: 29743376
- PMCID: PMC6052311
- DOI: 10.1128/JVI.00462-18
Transmembrane Protein pUL50 of Human Cytomegalovirus Inhibits ISGylation by Downregulating UBE1L
Abstract
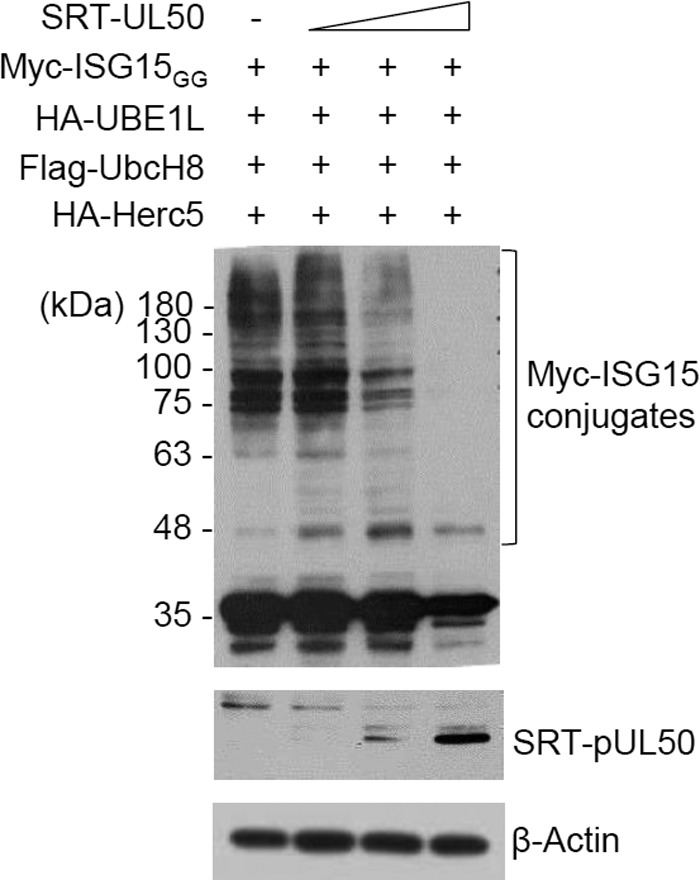
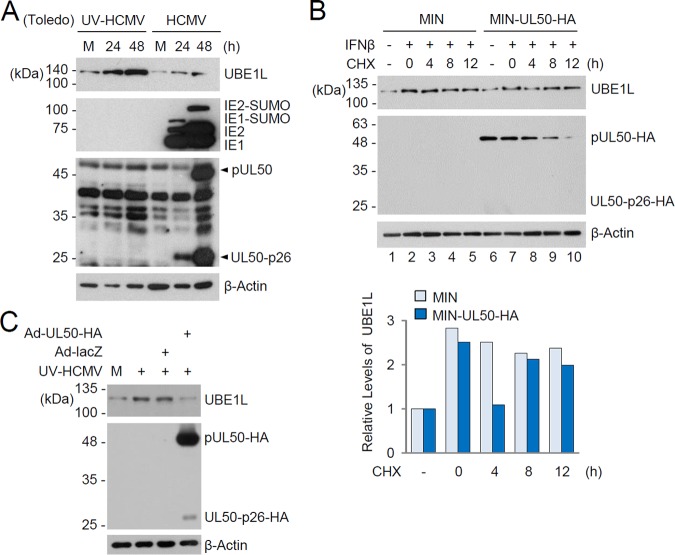
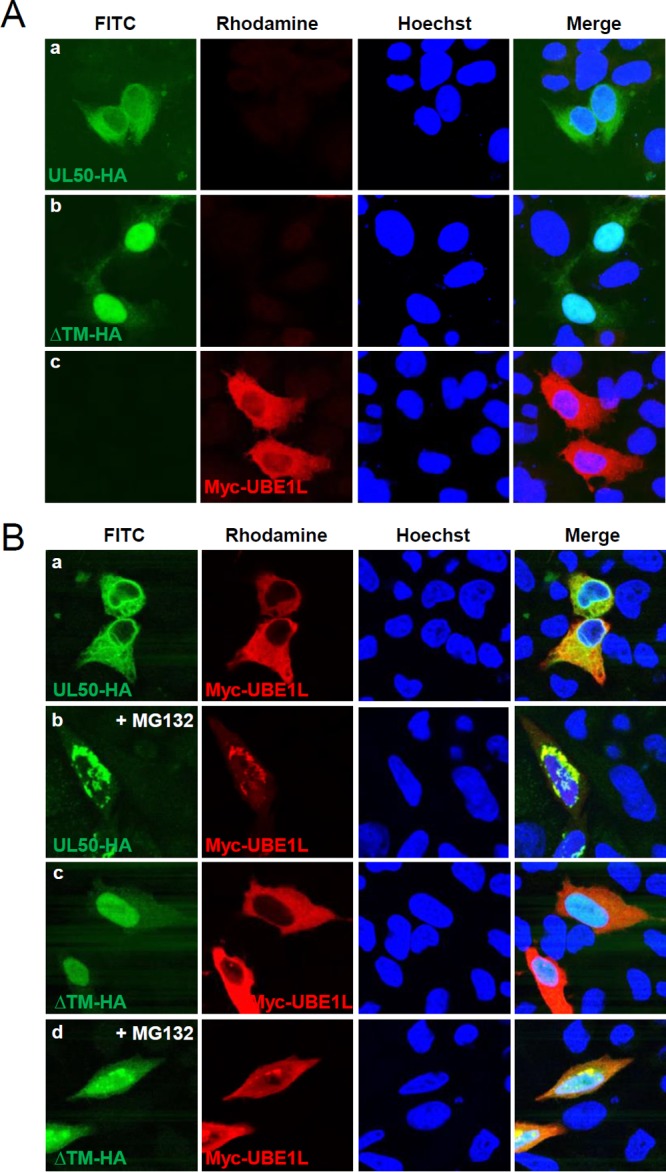
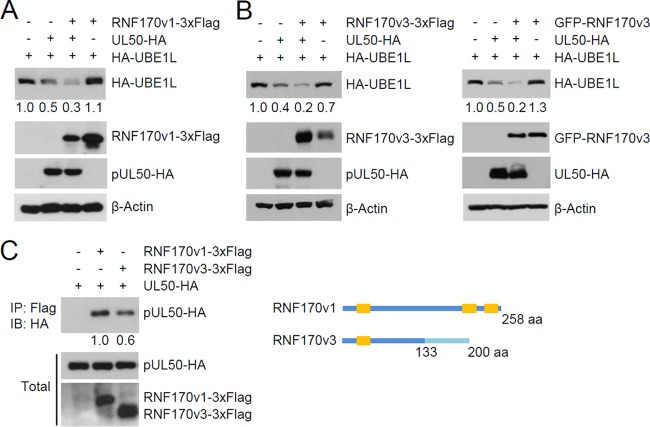
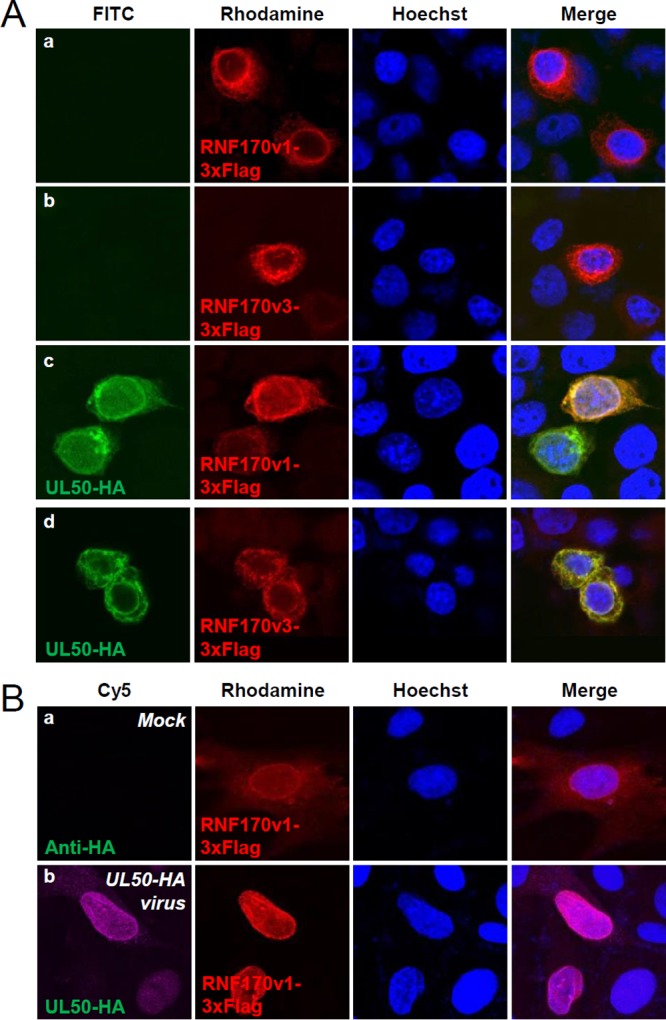
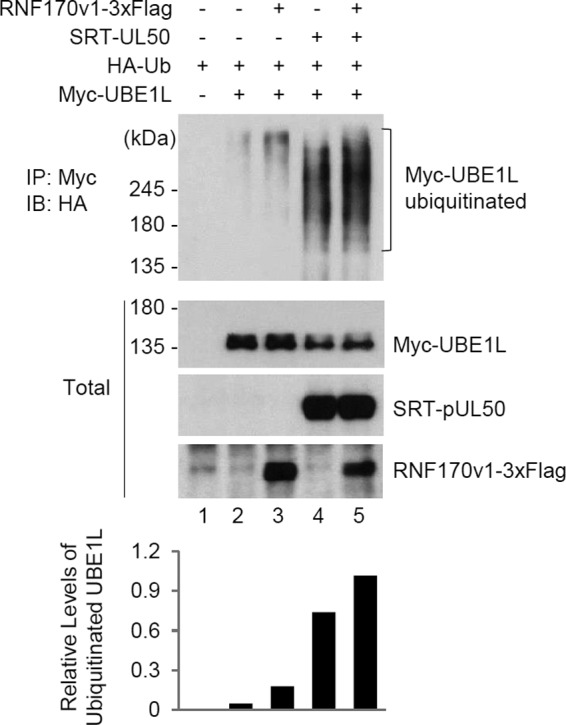
Interferon-stimulated gene 15 (ISG15) encodes a ubiquitin-like protein that can be conjugated to proteins via an enzymatic cascade involving the E1, E2, and E3 enzymes. ISG15 expression and protein ISGylation modulate viral infection; however, the viral mechanisms regulating the function of ISG15 and ISGylation are not well understood. We recently showed that ISGylation suppresses the growth of human cytomegalovirus (HCMV) at multiple steps of the virus life cycle and that the virus-encoded pUL26 protein inhibits protein ISGylation. In this study, we demonstrate that the HCMV UL50-encoded transmembrane protein, a component of the nuclear egress complex, also inhibits ISGylation. pUL50 interacted with UBE1L, an E1-activating enzyme for ISGylation, and (to a lesser extent) with ISG15, as did pUL26. However, unlike pUL26, pUL50 caused proteasomal degradation of UBE1L. The UBE1L level induced in human fibroblast cells by interferon beta treatment or virus infection was reduced by pUL50 expression. This activity of pUL50 involved the transmembrane (TM) domain within its C-terminal region, although pUL50 could interact with UBE1L in a manner independent of the TM domain. Consistently, colocalization of pUL50 with UBE1L was observed in cells treated with a proteasome inhibitor. Furthermore, we found that RNF170, an endoplasmic reticulum (ER)-associated ubiquitin E3 ligase, interacted with pUL50 and promoted pUL50-mediated UBE1L degradation via ubiquitination. Our results demonstrate a novel role for the pUL50 transmembrane protein of HCMV in the regulation of protein ISGylation.IMPORTANCE Proteins can be conjugated covalently by ubiquitin or ubiquitin-like proteins, such as SUMO and ISG15. ISG15 is highly induced in viral infection, and ISG15 conjugation, termed ISGylation, plays important regulatory roles in viral growth. Although ISGylation has been shown to negatively affect many viruses, including human cytomegalovirus (HCMV), viral countermeasures that might modulate ISGylation are not well understood. In the present study, we show that the transmembrane protein encoded by HCMV UL50 inhibits ISGylation by causing proteasomal degradation of UBE1L, an E1-activating enzyme for ISGylation. This pUL50 activity requires membrane targeting. In support of this finding, RNF170, an ER-associated ubiquitin E3 ligase, interacts with pUL50 and promotes UL50-mediated UBE1L ubiquitination and degradation. Our results provide the first evidence, to our knowledge, that viruses can regulate ISGylation by directly targeting the ISGylation E1 enzyme.
Keywords: ISGylation; UBE1L; UL50; cytomegalovirus.
Copyright © 2018 American Society for Microbiology.
Figures



Similar articles
-
The Human Cytomegalovirus Transmembrane Protein pUL50 Induces Loss of VCP/p97 and Is Regulated by a Small Isoform of pUL50.J Virol. 2020 Jun 16;94(13):e00110-20. doi: 10.1128/JVI.00110-20. Print 2020 Jun 16. J Virol. 2020. PMID: 32321808 Free PMC article.
-
Consecutive Inhibition of ISG15 Expression and ISGylation by Cytomegalovirus Regulators.PLoS Pathog. 2016 Aug 26;12(8):e1005850. doi: 10.1371/journal.ppat.1005850. eCollection 2016 Aug. PLoS Pathog. 2016. PMID: 27564865 Free PMC article.
-
The Abundant Tegument Protein pUL25 of Human Cytomegalovirus Prevents Proteasomal Degradation of pUL26 and Supports Its Suppression of ISGylation.J Virol. 2018 Nov 27;92(24):e01180-18. doi: 10.1128/JVI.01180-18. Print 2018 Dec 15. J Virol. 2018. PMID: 30282718 Free PMC article.
-
Identifying the ERAD ubiquitin E3 ligases for viral and cellular targeting of MHC class I.Mol Immunol. 2015 Dec;68(2 Pt A):106-11. doi: 10.1016/j.molimm.2015.07.005. Epub 2015 Jul 22. Mol Immunol. 2015. PMID: 26210183 Free PMC article. Review.
-
[Advances in ubiquitin-like protein ISG15-mediated anti-viral response].Bing Du Xue Bao. 2012 Jan;28(1):78-83. Bing Du Xue Bao. 2012. PMID: 22416355 Review. Chinese.
Cited by
-
Innate Immune Responses to Herpesvirus Infection.Cells. 2021 Aug 18;10(8):2122. doi: 10.3390/cells10082122. Cells. 2021. PMID: 34440891 Free PMC article. Review.
-
'Come together'-The Regulatory Interaction of Herpesviral Nuclear Egress Proteins Comprises Both Essential and Accessory Functions.Cells. 2022 Jun 4;11(11):1837. doi: 10.3390/cells11111837. Cells. 2022. PMID: 35681532 Free PMC article. Review.
-
Post-translational Control of Innate Immune Signaling Pathways by Herpesviruses.Front Microbiol. 2019 Nov 14;10:2647. doi: 10.3389/fmicb.2019.02647. eCollection 2019. Front Microbiol. 2019. PMID: 31798565 Free PMC article. Review.
-
Insights into the Transcriptome of Human Cytomegalovirus: A Comprehensive Review.Viruses. 2023 Aug 8;15(8):1703. doi: 10.3390/v15081703. Viruses. 2023. PMID: 37632045 Free PMC article. Review.
-
The battle between host antiviral innate immunity and immune evasion by cytomegalovirus.Cell Mol Life Sci. 2024 Aug 9;81(1):341. doi: 10.1007/s00018-024-05369-y. Cell Mol Life Sci. 2024. PMID: 39120730 Free PMC article. Review.
References
-
- Haas AL, Ahrens P, Bright PM, Ankel H. 1987. Interferon induces a 15-kilodalton protein exhibiting marked homology to ubiquitin. J Biol Chem 262:11315–11323. - PubMed
-
- Loeb KR, Haas AL. 1992. The interferon-inducible 15-kDa ubiquitin homolog conjugates to intracellular proteins. J Biol Chem 267:7806–7813. - PubMed
-
- Arimoto KI, Lochte S, Stoner SA, Burkart C, Zhang Y, Miyauchi S, Wilmes S, Fan JB, Heinisch JJ, Li Z, Yan M, Pellegrini S, Colland F, Piehler J, Zhang DE. 2017. STAT2 is an essential adaptor in USP18-mediated suppression of type I interferon signaling. Nat Struct Mol Biol 24:279–289. doi:10.1038/nsmb.3378. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
