

Chemical synapses without synaptic vesicles: Purinergic neurotransmission through a CALHM1 channel-mitochondrial signaling complex
- PMID: 29739879
- PMCID: PMC5966022
- DOI: 10.1126/scisignal.aao1815
Chemical synapses without synaptic vesicles: Purinergic neurotransmission through a CALHM1 channel-mitochondrial signaling complex
Abstract
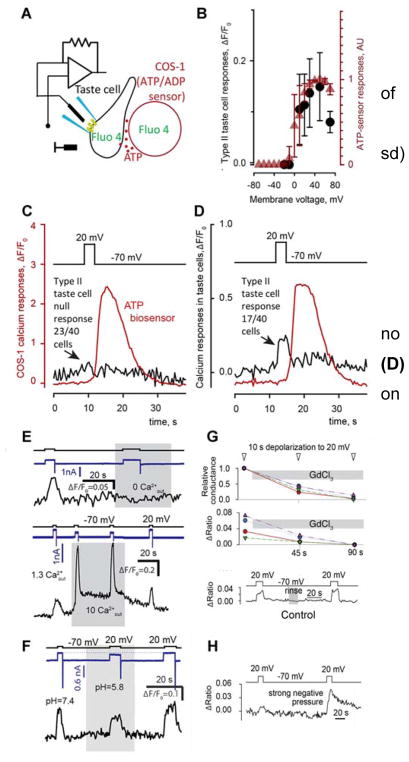
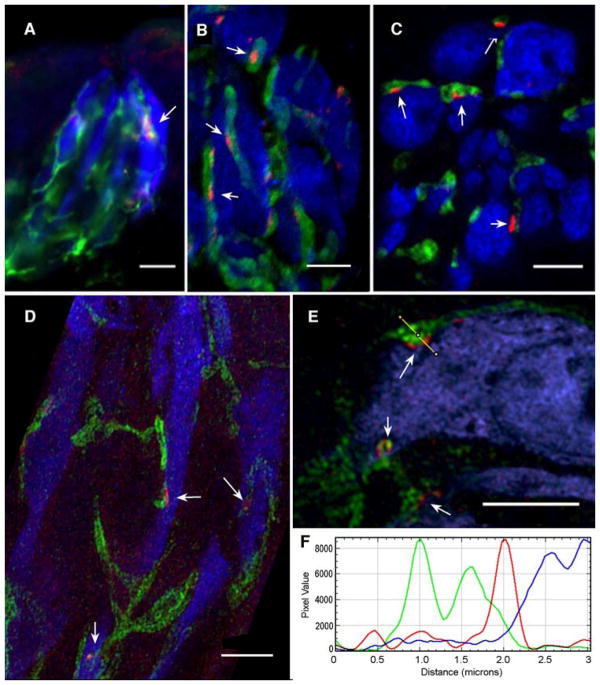
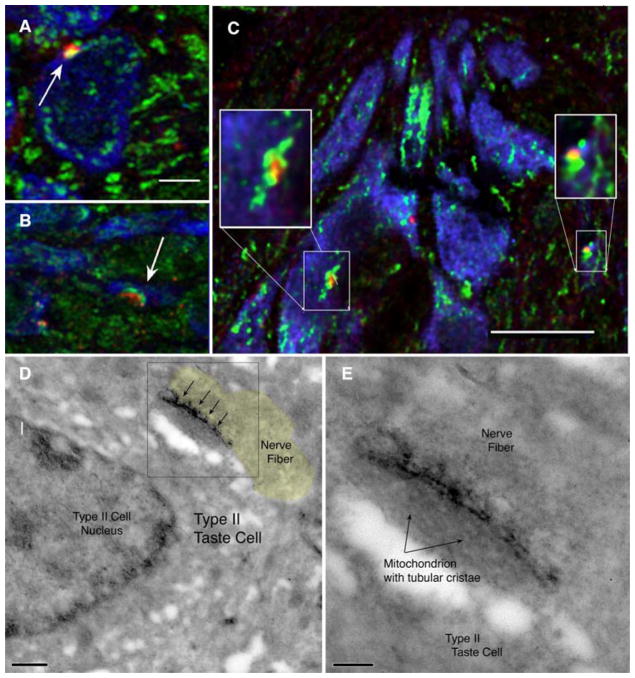
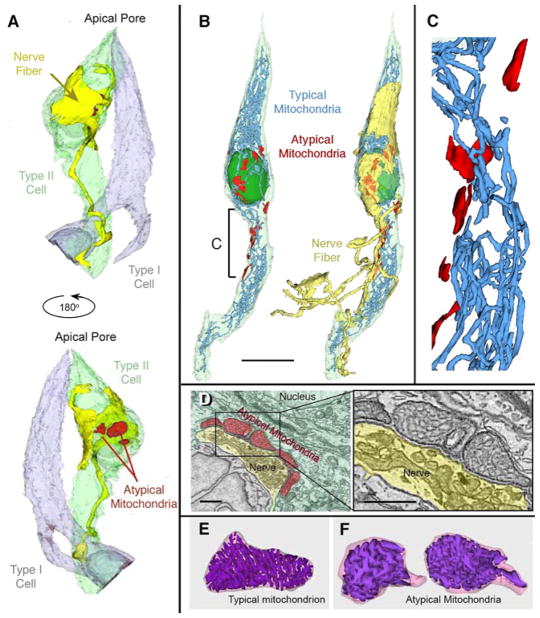
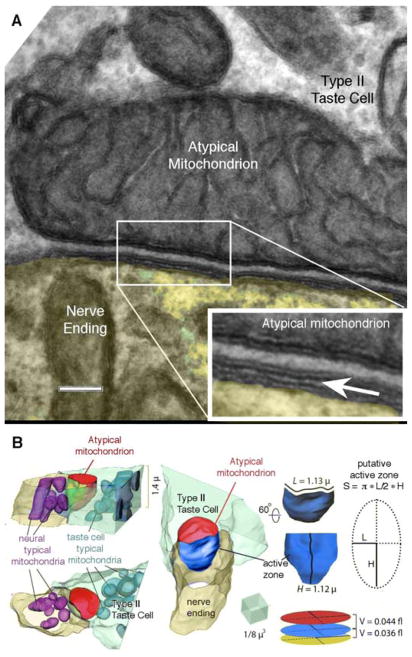
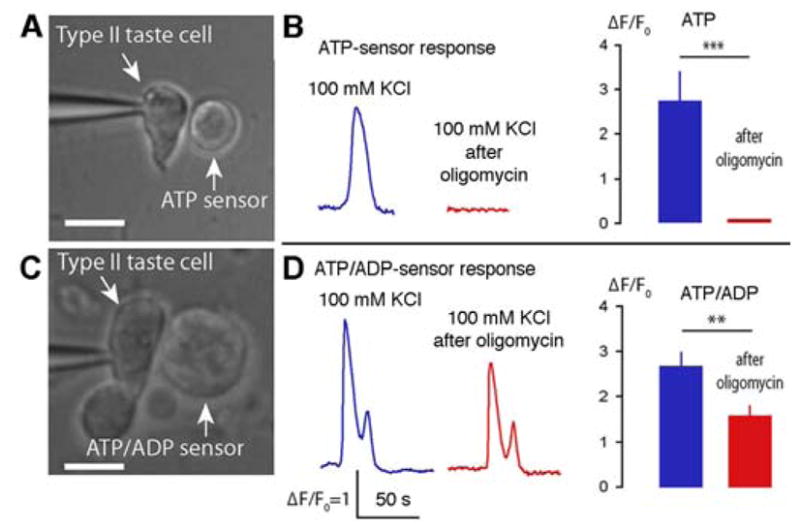
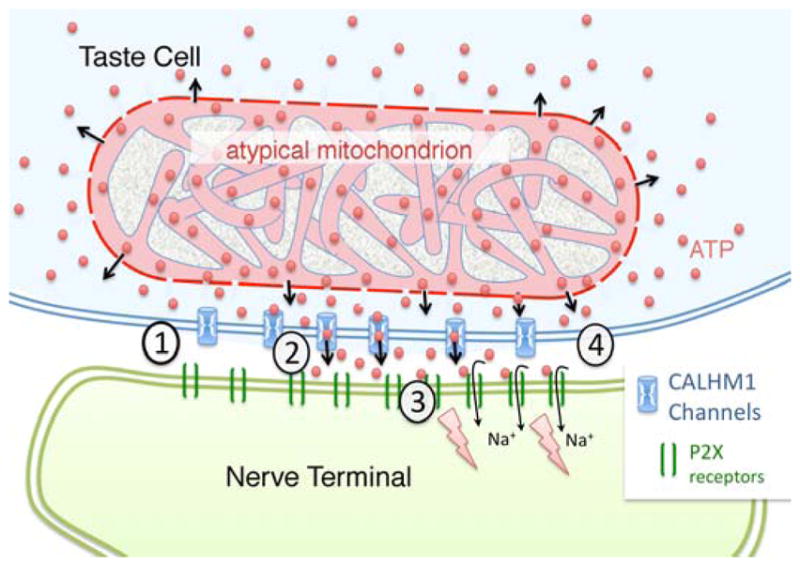
Conventional chemical synapses in the nervous system involve a presynaptic accumulation of neurotransmitter-containing vesicles, which fuse with the plasma membrane to release neurotransmitters that activate postsynaptic receptors. In taste buds, type II receptor cells do not have conventional synaptic features but nonetheless show regulated release of their afferent neurotransmitter, ATP, through a large-pore, voltage-gated channel, CALHM1. Immunohistochemistry revealed that CALHM1 was localized to points of contact between the receptor cells and sensory nerve fibers. Ultrastructural and super-resolution light microscopy showed that the CALHM1 channels were consistently associated with distinctive, large (1- to 2-μm) mitochondria spaced 20 to 40 nm from the presynaptic membrane. Pharmacological disruption of the mitochondrial respiratory chain limited the ability of taste cells to release ATP, suggesting that the immediate source of released ATP was the mitochondrion rather than a cytoplasmic pool of ATP. These large mitochondria may serve as both a reservoir of releasable ATP and the site of synthesis. The juxtaposition of the large mitochondria to areas of membrane displaying CALHM1 also defines a restricted compartment that limits the influx of Ca2+ upon opening of the nonselective CALHM1 channels. These findings reveal a distinctive organelle signature and functional organization for regulated, focal release of purinergic signals in the absence of synaptic vesicles.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Similar articles
-
How do taste cells lacking synapses mediate neurotransmission? CALHM1, a voltage-gated ATP channel.Bioessays. 2013 Dec;35(12):1111-8. doi: 10.1002/bies.201300077. Epub 2013 Sep 17. Bioessays. 2013. PMID: 24105910 Free PMC article. Review.
-
CALHM1 ion channel mediates purinergic neurotransmission of sweet, bitter and umami tastes.Nature. 2013 Mar 14;495(7440):223-6. doi: 10.1038/nature11906. Epub 2013 Mar 6. Nature. 2013. PMID: 23467090 Free PMC article.
-
Ultrastructural localization of calcium homeostasis modulator 1 in mouse taste buds.Chem Senses. 2024 Jan 1;49:bjae019. doi: 10.1093/chemse/bjae019. Chem Senses. 2024. PMID: 38761122
-
Action potentials and ion conductances in wild-type and CALHM1-knockout type II taste cells.J Neurophysiol. 2017 May 1;117(5):1865-1876. doi: 10.1152/jn.00835.2016. Epub 2017 Feb 15. J Neurophysiol. 2017. PMID: 28202574 Free PMC article.
-
Calcium homeostasis modulator (CALHM) ion channels.Pflugers Arch. 2016 Mar;468(3):395-403. doi: 10.1007/s00424-015-1757-6. Epub 2015 Nov 25. Pflugers Arch. 2016. PMID: 26603282 Free PMC article. Review.
Cited by
-
Give-and-take of gustation: the interplay between gustatory neurons and taste buds.Chem Senses. 2024 Jan 1;49:bjae029. doi: 10.1093/chemse/bjae029. Chem Senses. 2024. PMID: 39078723 Review.
-
Sensing Senses: Optical Biosensors to Study Gustation.Sensors (Basel). 2020 Mar 25;20(7):1811. doi: 10.3390/s20071811. Sensors (Basel). 2020. PMID: 32218129 Free PMC article. Review.
-
Interkingdom Detection of Bacterial Quorum-Sensing Molecules by Mammalian Taste Receptors.Microorganisms. 2023 May 16;11(5):1295. doi: 10.3390/microorganisms11051295. Microorganisms. 2023. PMID: 37317269 Free PMC article. Review.
-
Peripheral and central mechanisms of cough hypersensitivity.J Thorac Dis. 2020 Sep;12(9):5179-5193. doi: 10.21037/jtd-2020-icc-007. J Thorac Dis. 2020. PMID: 33145095 Free PMC article. Review.
-
Sour taste: receptors, cells and circuits.Curr Opin Physiol. 2021 Apr;20:8-15. doi: 10.1016/j.cophys.2020.12.006. Epub 2021 Jan 13. Curr Opin Physiol. 2021. PMID: 33709046 Free PMC article.
References
-
- Foster M, Sherrington SC. A Textbook of Physiology, Part 3. 7. Macmillan; London: 1897.
-
- Grakoui A, Bromley SK, Sumen C, Davis MM, Shaw AS, Allen PM, Dustin ML. The immunological synapse: a molecular machine controlling T cell activation. Science. 1999;285:221. published online EpubJul 09. - PubMed
-
- Vandenbeuch A, Larson ED, Anderson CB, Smith SA, Ford AP, Finger TE, Kinnamon SC. Postsynaptic P2X3-containing receptors in gustatory nerve fibres mediate responses to all taste qualities in mice. J Physiol. 2015;593:1113. doi: 10.1113/jphysiol.2014.281014. published online EpubMar 1. - DOI - PMC - PubMed
-
- Vandenbeuch A, Anderson CB, Ford AP, Smith S, Finger TE, Kinnamon SC. A selective P2X3, P2X2/3 receptor antagonist abolishes responses to all taste stimuli in mice. Chem Senses. 2013;38:86. doi: 10.1093/chemse/bjt036). - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
