

Effects of hypervariable regions in spike protein on pathogenicity, tropism, and serotypes of infectious bronchitis virus
- PMID: 29684409
- PMCID: PMC7114591
- DOI: 10.1016/j.virusres.2018.04.013
Effects of hypervariable regions in spike protein on pathogenicity, tropism, and serotypes of infectious bronchitis virus
Abstract
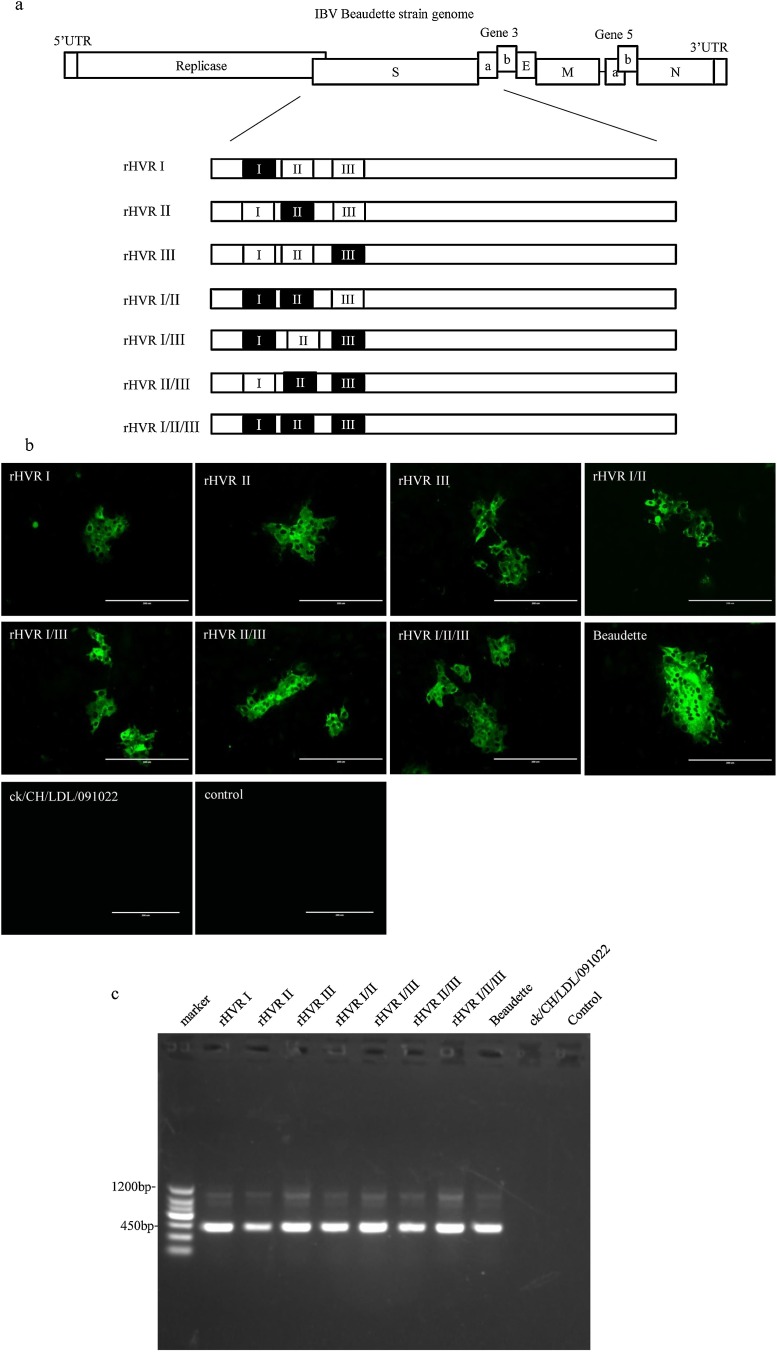
To study the roles of hypervariable regions (HVRs) in receptor-binding subunit S1 of the spike protein, we manipulated the genome of the IBV Beaudette strain using a reverse genetics system to construct seven recombinant strains by separately or simultaneously replacing the three HVRs of the Beaudette strain with the corresponding fragments from a QX-like nephropathogenic isolate ck/CH/LDL/091022 from China. We characterized the growth properties of these recombinant IBVs in Vero cells and embryonated eggs, and their pathogenicity, tropism, and serotypes in specific pathogen-free (SPF) chickens. All seven recombinant IBVs proliferated in Vero cells, but the heterogenous HVRs could reduce their capacity for adsorption during in vitro infection. The recombinant IBVs did not significantly increase the pathogenicity compared with the Beaudette strain in SPF chickens, and they still shared the same serotype as the Beaudette strain, but the antigenic relatedness values between the recombinant strain and Beaudette strain generally decreased with the increase in the number of the HVRs exchanged. The results of this study demonstrate the functions of HVRs and they may help to develop a vaccine candidate, as well as providing insights into the prevention and control of IBV.
Keywords: Hypervariable region; Infectious bronchitis virus; Infectious clone; Recombinant IBV; Reverse genetic vaccine.
Copyright © 2018 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
The S2 Subunit of Infectious Bronchitis Virus Beaudette Is a Determinant of Cellular Tropism.J Virol. 2018 Sep 12;92(19):e01044-18. doi: 10.1128/JVI.01044-18. Print 2018 Oct 1. J Virol. 2018. PMID: 30021894 Free PMC article.
-
Characterization of the complete genome, antigenicity, pathogenicity, tissue tropism, and shedding of a recombinant avian infectious bronchitis virus with a ck/CH/LJL/140901-like backbone and an S2 fragment from a 4/91-like virus.Virus Res. 2018 Jan 15;244:99-109. doi: 10.1016/j.virusres.2017.11.007. Epub 2017 Nov 12. Virus Res. 2018. PMID: 29141204 Free PMC article.
-
Genetics, antigenicity and virulence properties of three infectious bronchitis viruses isolated from a single tracheal sample in a chicken with respiratory problems.Virus Res. 2018 Sep 15;257:82-93. doi: 10.1016/j.virusres.2018.09.009. Epub 2018 Sep 18. Virus Res. 2018. PMID: 30240807 Free PMC article.
-
The avian coronavirus spike protein.Virus Res. 2014 Dec 19;194:37-48. doi: 10.1016/j.virusres.2014.10.009. Epub 2014 Oct 17. Virus Res. 2014. PMID: 25451062 Free PMC article. Review.
-
Current situation, genetic relationship and control measures of infectious bronchitis virus variants circulating in African regions.J Basic Appl Zool. 2016 Aug;76:20-30. doi: 10.1016/j.jobaz.2016.08.002. Epub 2016 Nov 20. J Basic Appl Zool. 2016. PMID: 32288944 Free PMC article. Review.
Cited by
-
A multiplex real-time reverse transcription polymerase chain reaction assay for differentiation of classical and variant II strains of avian infectious bronchitis virus.Arch Virol. 2022 Dec;167(12):2729-2741. doi: 10.1007/s00705-022-05603-7. Epub 2022 Sep 29. Arch Virol. 2022. PMID: 36175795 Free PMC article.
-
Avian infectious bronchitis virus (AIBV) review by continent.Front Cell Infect Microbiol. 2024 Feb 5;14:1325346. doi: 10.3389/fcimb.2024.1325346. eCollection 2024. Front Cell Infect Microbiol. 2024. PMID: 38375362 Free PMC article. Review.
-
Genetic and biological characteristics of four novel recombinant avian infectious bronchitis viruses isolated in China.Virus Res. 2019 Apr 2;263:87-97. doi: 10.1016/j.virusres.2019.01.007. Epub 2019 Jan 11. Virus Res. 2019. PMID: 30641197 Free PMC article.
-
Recombinant infectious bronchitis coronavirus H120 with the spike protein S1 gene of the nephropathogenic IBYZ strain remains attenuated but induces protective immunity.Vaccine. 2020 Mar 30;38(15):3157-3168. doi: 10.1016/j.vaccine.2020.01.001. Epub 2020 Feb 11. Vaccine. 2020. PMID: 32057575 Free PMC article.
-
Antigenic and Pathogenic Characteristics of QX-Type Avian Infectious Bronchitis Virus Strains Isolated in Southwestern China.Viruses. 2019 Dec 13;11(12):1154. doi: 10.3390/v11121154. Viruses. 2019. PMID: 31847269 Free PMC article.
References
-
- Beato M.S., De Battisti C., Terregino C., Drago A., Capua I., Ortali G. Evidence of circulation of a Chinese strain of infectious bronchitis virus (QXIBV) in Italy. Vet. Rec. 2005;156(May (22)):720. - PubMed
-
- Bochkov Y.A., Batchenko G.V., Shcherbakova L.O., Borisov A.V., Drygin V.V. Molecular epizootiology of avian infectious bronchitis in Russia. Avian Pathol. 2006;35(5):379–393. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
