

The G-Protein-Coupled Receptor ALX/Fpr2 Regulates Adaptive Immune Responses in Mouse Submandibular Glands
- PMID: 29684359
- PMCID: PMC6024187
- DOI: 10.1016/j.ajpath.2018.04.003
The G-Protein-Coupled Receptor ALX/Fpr2 Regulates Adaptive Immune Responses in Mouse Submandibular Glands
Abstract
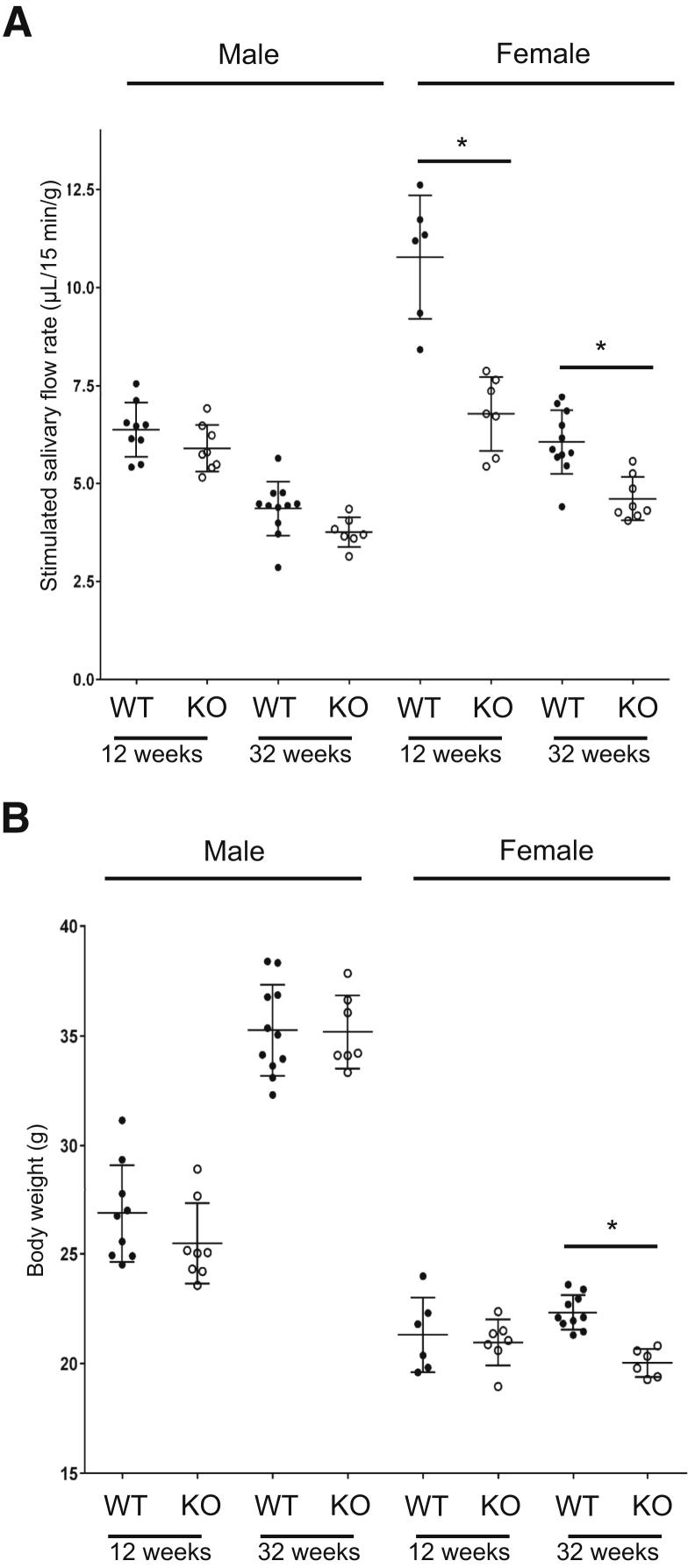
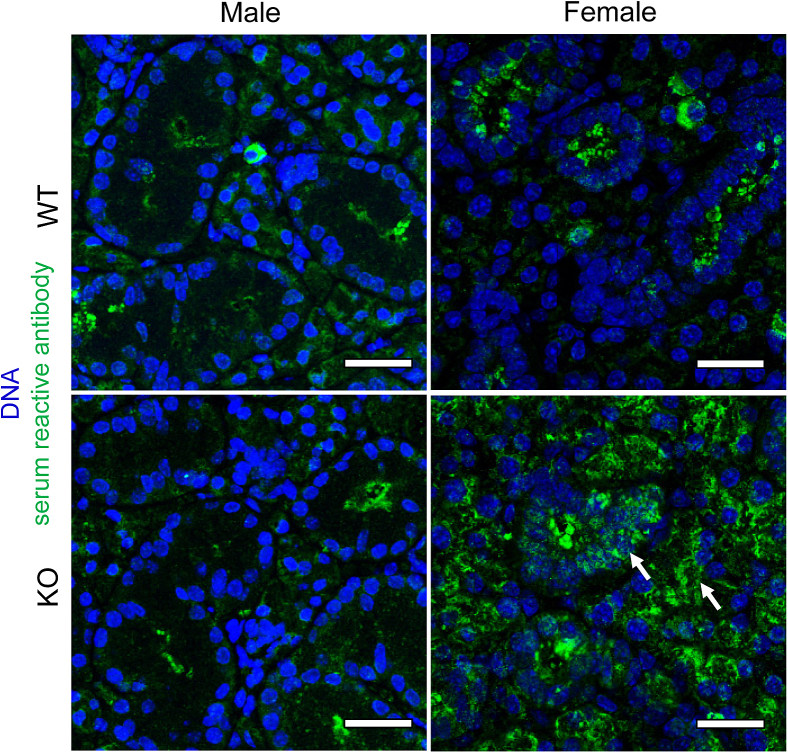
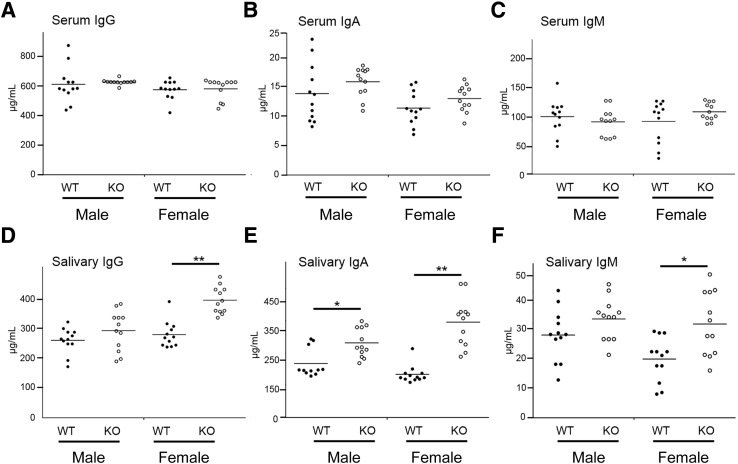
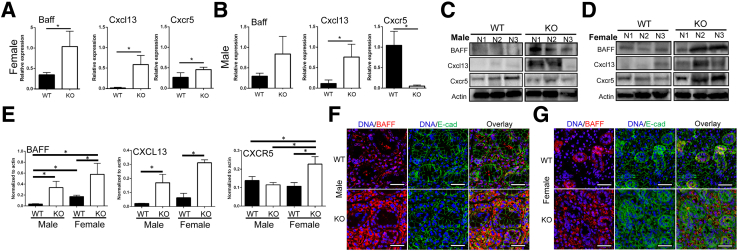
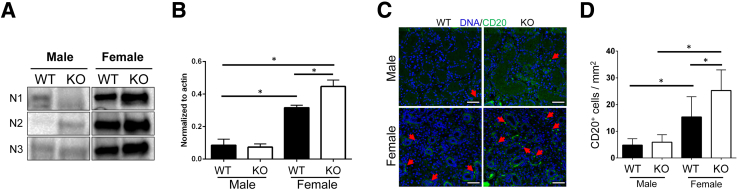
Lipoxin receptor (ALX)/N-formyl peptide receptor (FPR)-2 is a G-protein-coupled receptor that has multiple binding partners, including the endogenous lipid mediators resolvin D1, lipoxin A4, and the Ca2+-dependent phospholipid-binding protein annexin A1. Previous studies have demonstrated that resolvin D1 activates ALX/Fpr2 to resolve salivary gland inflammation in the NOD/ShiLtJ mouse model of Sjögren syndrome. Moreover, mice lacking the ALX/Fpr2 display an exacerbated salivary gland inflammation in response to lipopolysaccharide. Additionally, activation of ALX/Fpr2 has been shown to be important for regulating antibody production in B cells. These previous studies indicate that ALX/Fpr2 promotes resolution of salivary gland inflammation while modulating adaptive immunity, suggesting the need for investigation of the role of ALX/Fpr2 in regulating antibody production and secretory function in mouse salivary glands. Our results indicate that aging female knockout mice lacking ALX/Fpr2 display a significant reduction in saliva flow rates and weight loss, an increased expression of autoimmune-associated genes, an up-regulation of autoantibody production, and increased CD20-positive B-cell population. Although not all effects were noted among the male knockout mice, the results nonetheless indicate that ALX/Fpr2 is clearly involved in the adaptive immunity and secretory function in salivary glands, with further investigation warranted to determine the cause(s) of these between-sex differences.
Copyright © 2018 American Society for Investigative Pathology. Published by Elsevier Inc. All rights reserved.
Figures
Similar articles
-
ALX/FPR2 Modulates Anti-Inflammatory Responses in Mouse Submandibular Gland.Sci Rep. 2016 Apr 11;6:24244. doi: 10.1038/srep24244. Sci Rep. 2016. PMID: 27064029 Free PMC article.
-
Sex-mediated elevation of the specialized pro-resolving lipid mediator levels in a Sjögren's syndrome mouse model.FASEB J. 2020 Jun;34(6):7733-7744. doi: 10.1096/fj.201902196R. Epub 2020 Apr 11. FASEB J. 2020. PMID: 32277856 Free PMC article.
-
ALX/FPR2 receptor for RvD1 is expressed and functional in salivary glands.Am J Physiol Cell Physiol. 2014 Jan 15;306(2):C178-85. doi: 10.1152/ajpcell.00284.2013. Epub 2013 Nov 20. Am J Physiol Cell Physiol. 2014. PMID: 24259417 Free PMC article.
-
Treating neutrophilic inflammation in COPD by targeting ALX/FPR2 resolution pathways.Pharmacol Ther. 2013 Dec;140(3):280-9. doi: 10.1016/j.pharmthera.2013.07.007. Epub 2013 Jul 21. Pharmacol Ther. 2013. PMID: 23880288 Review.
-
Recent advances in the design and development of formyl peptide receptor 2 (FPR2/ALX) agonists as pro-resolving agents with diverse therapeutic potential.Eur J Med Chem. 2021 Mar 5;213:113167. doi: 10.1016/j.ejmech.2021.113167. Epub 2021 Jan 12. Eur J Med Chem. 2021. PMID: 33486199 Review.
Cited by
-
Formyl peptide receptors in the mucosal immune system.Exp Mol Med. 2020 Oct;52(10):1694-1704. doi: 10.1038/s12276-020-00518-2. Epub 2020 Oct 20. Exp Mol Med. 2020. PMID: 33082511 Free PMC article. Review.
-
Aspirin Triggered Resolvin D1 reduces inflammation and restores saliva secretion in a Sjögren's syndrome mouse model.Rheumatology (Oxford). 2019 Jul 1;58(7):1285-1292. doi: 10.1093/rheumatology/kez072. Rheumatology (Oxford). 2019. PMID: 30877775 Free PMC article.
-
Specialized pro-resolving receptors are expressed in salivary glands with Sjögren's syndrome.Ann Diagn Pathol. 2022 Feb;56:151865. doi: 10.1016/j.anndiagpath.2021.151865. Epub 2021 Nov 17. Ann Diagn Pathol. 2022. PMID: 34847389 Free PMC article.
-
Lipoxin alleviates oxidative stress: a state-of-the-art review.Inflamm Res. 2022 Nov;71(10-11):1169-1179. doi: 10.1007/s00011-022-01621-y. Inflamm Res. 2022. PMID: 35947143 Review.
-
A combination treatment of low-dose dexamethasone and aspirin-triggered resolvin D1 reduces Sjögren syndrome-like features in a mouse model.JADA Found Sci. 2023;2:100016. doi: 10.1016/j.jfscie.2022.100016. Epub 2022 Nov 17. JADA Found Sci. 2023. PMID: 37622089 Free PMC article.
References
-
- Fredman G., Serhan C.N. Specialized pro-resolving mediators: wiring the circuitry of effector immune and tissue homeostasis. Endod Top. 2011;24:39–58.
-
- Ariel A., Serhan C.N. Resolvins and protectins in the termination program of acute inflammation. Trends Immunol. 2007;28:176–183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
