

Palmitoleic acid (16:1n7) increases oxygen consumption, fatty acid oxidation and ATP content in white adipocytes
- PMID: 29554895
- PMCID: PMC5859716
- DOI: 10.1186/s12944-018-0710-z
Palmitoleic acid (16:1n7) increases oxygen consumption, fatty acid oxidation and ATP content in white adipocytes
Abstract
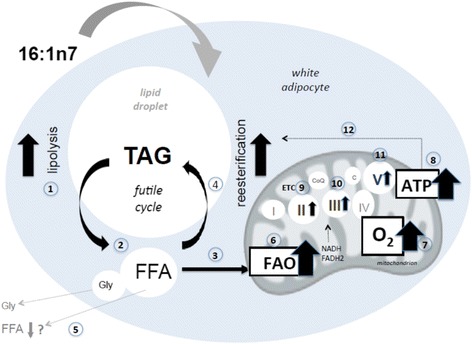
Background: We have recently demonstrated that palmitoleic acid (16:1n7) increases lipolysis, glucose uptake and glucose utilization for energy production in white adipose cells. In the present study, we tested the hypothesis that palmitoleic acid modulates bioenergetic activity in white adipocytes.
Methods: For this, 3 T3-L1 pre-adipocytes were differentiated into mature adipocytes in the presence (or absence) of palmitic (16:0) or palmitoleic (16:1n7) acid at 100 or 200 μM. The following parameters were evaluated: lipolysis, lipogenesis, fatty acid (FA) oxidation, ATP content, oxygen consumption, mitochondrial mass, citrate synthase activity and protein content of mitochondrial oxidative phosphorylation (OXPHOS) complexes.
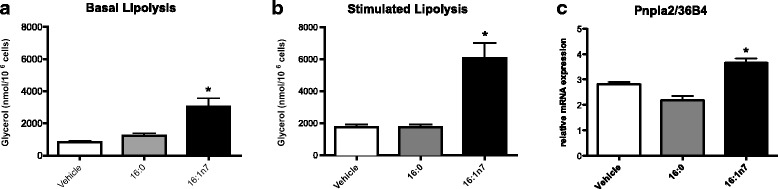
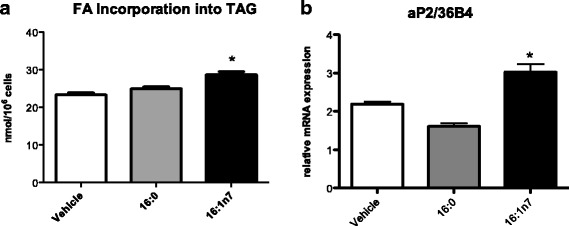
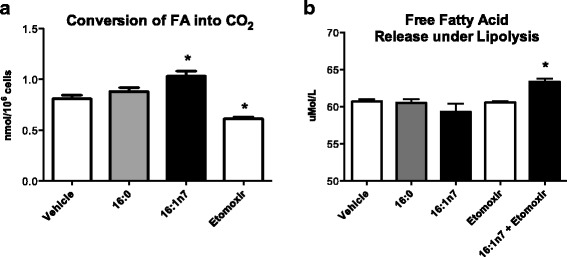
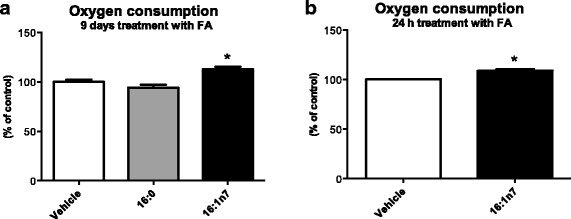
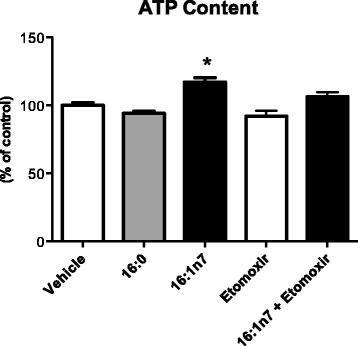
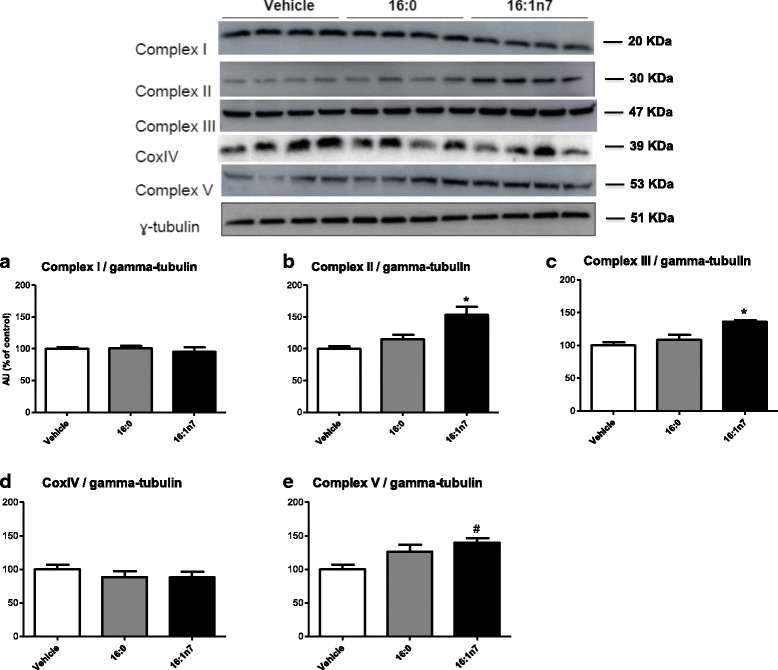
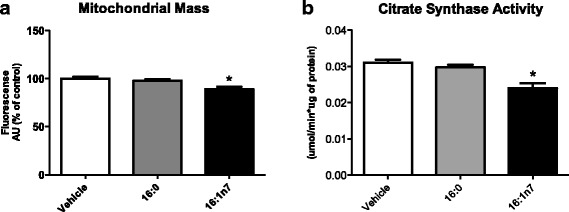
Results: Treatment with 16:1n7 during 9 days raised basal and isoproterenol-stimulated lipolysis, FA incorporation into triacylglycerol (TAG), FA oxidation, oxygen consumption, protein expression of subunits representing OXPHOS complex II, III, and V and intracellular ATP content. These effects were not observed in adipocytes treated with 16:0.
Conclusions: Palmitoleic acid, by concerted action on lipolysis, FA esterification, mitochondrial FA oxidation, oxygen consumption and ATP content, does enhance white adipocyte energy expenditure and may act as local hormone.
Keywords: Beta-oxidation; Bioenergetics; Lipogenesis; Lipolysis; Mitochondria; Triglyceride/fatty acid cycle.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
Palmitoleic acid (n-7) increases white adipocyte lipolysis and lipase content in a PPARα-dependent manner.Am J Physiol Endocrinol Metab. 2013 Nov 1;305(9):E1093-102. doi: 10.1152/ajpendo.00082.2013. Epub 2013 Sep 10. Am J Physiol Endocrinol Metab. 2013. PMID: 24022867
-
Palmitoleic Acid Decreases Non-alcoholic Hepatic Steatosis and Increases Lipogenesis and Fatty Acid Oxidation in Adipose Tissue From Obese Mice.Front Endocrinol (Lausanne). 2020 Sep 30;11:537061. doi: 10.3389/fendo.2020.537061. eCollection 2020. Front Endocrinol (Lausanne). 2020. PMID: 33117273 Free PMC article.
-
Palmitoleic acid (n-7) increases white adipocytes GLUT4 content and glucose uptake in association with AMPK activation.Lipids Health Dis. 2014 Dec 20;13:199. doi: 10.1186/1476-511X-13-199. Lipids Health Dis. 2014. PMID: 25528561 Free PMC article.
-
[THE EXCESS OF PALMITIC FATTY ACID IN FOOD AS MAIN CAUSE OF LIPOIDOSIS OF INSULIN-DEPENDENT CELLS: SKELETAL MYOCYTES, CARDIO-MYOCYTES, PERIPORTAL HEPATOCYTES, KUPFFER MACROPHAGES AND B-CELLS OF PANCREAS].Klin Lab Diagn. 2016 Feb;61(2):68-77. Klin Lab Diagn. 2016. PMID: 27455558 Review. Russian.
-
The skinny on fat: lipolysis and fatty acid utilization in adipocytes.Trends Endocrinol Metab. 2009 Nov;20(9):424-8. doi: 10.1016/j.tem.2009.06.002. Epub 2009 Sep 30. Trends Endocrinol Metab. 2009. PMID: 19796963 Free PMC article. Review.
Cited by
-
Role of Exendin-4 in Brain Insulin Resistance, Mitochondrial Function, and Neurite Outgrowth in Neurons under Palmitic Acid-Induced Oxidative Stress.Antioxidants (Basel). 2021 Jan 9;10(1):78. doi: 10.3390/antiox10010078. Antioxidants (Basel). 2021. PMID: 33435277 Free PMC article.
-
Oleic and Linoleic Acids Induce the Release of Neutrophil Extracellular Traps via Pannexin 1-Dependent ATP Release and P2X1 Receptor Activation.Front Vet Sci. 2020 Jun 5;7:260. doi: 10.3389/fvets.2020.00260. eCollection 2020. Front Vet Sci. 2020. PMID: 32582772 Free PMC article.
-
The Regulation of Lipokines by Environmental Factors.Nutrients. 2019 Oct 11;11(10):2422. doi: 10.3390/nu11102422. Nutrients. 2019. PMID: 31614481 Free PMC article. Review.
-
Preclinical modeling of metabolic syndrome to study the pleiotropic effects of novel antidiabetic therapy independent of obesity.Sci Rep. 2024 Sep 5;14(1):20665. doi: 10.1038/s41598-024-71202-y. Sci Rep. 2024. PMID: 39237601 Free PMC article.
-
The molecular and metabolic program by which white adipocytes adapt to cool physiologic temperatures.PLoS Biol. 2021 May 12;19(5):e3000988. doi: 10.1371/journal.pbio.3000988. eCollection 2021 May. PLoS Biol. 2021. PMID: 33979328 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
