

Intrahost Norovirus Evolution in Chronic Infection Over 5 Years of Shedding in a Kidney Transplant Recipient
- PMID: 29552005
- PMCID: PMC5840165
- DOI: 10.3389/fmicb.2018.00371
Intrahost Norovirus Evolution in Chronic Infection Over 5 Years of Shedding in a Kidney Transplant Recipient
Abstract
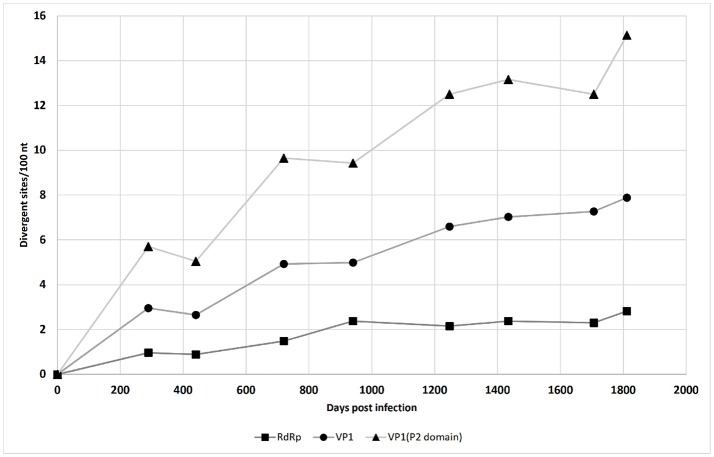
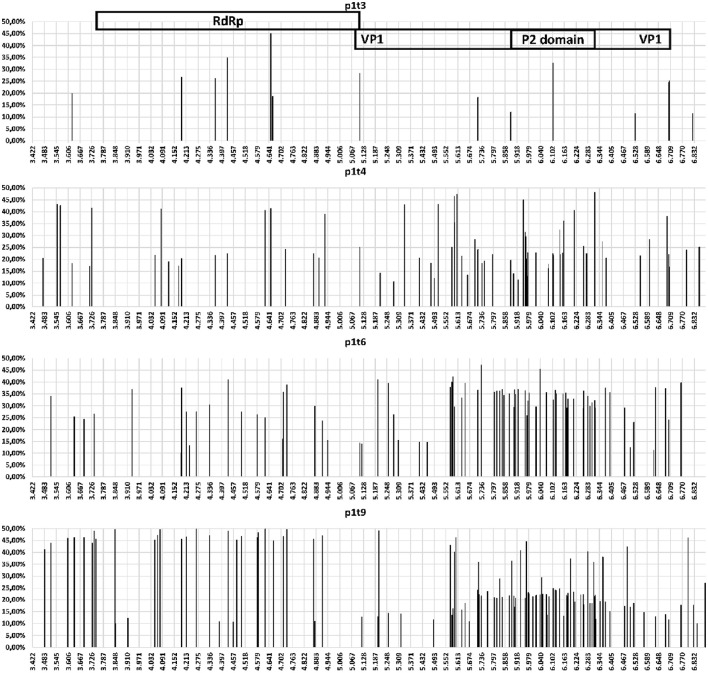
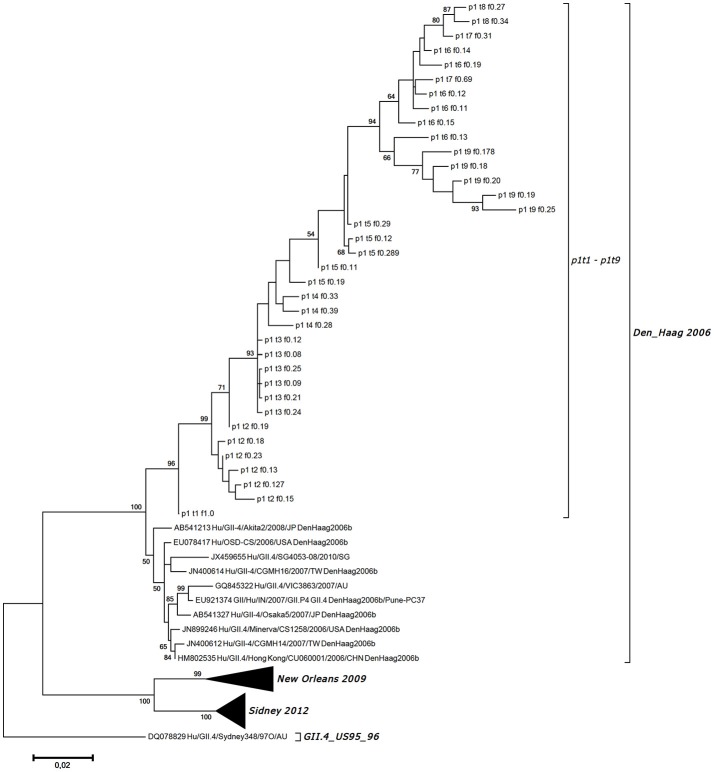
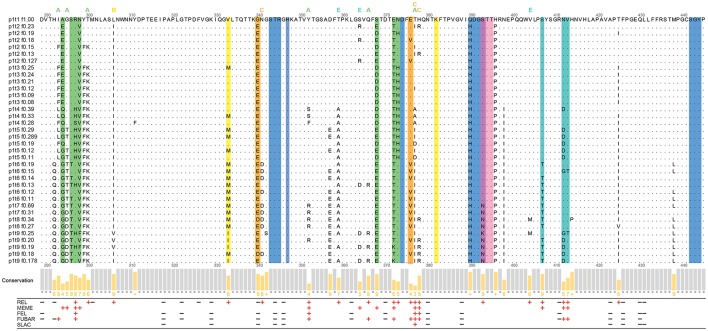
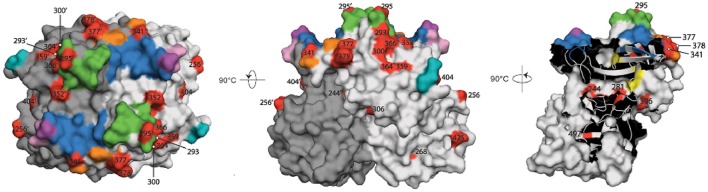
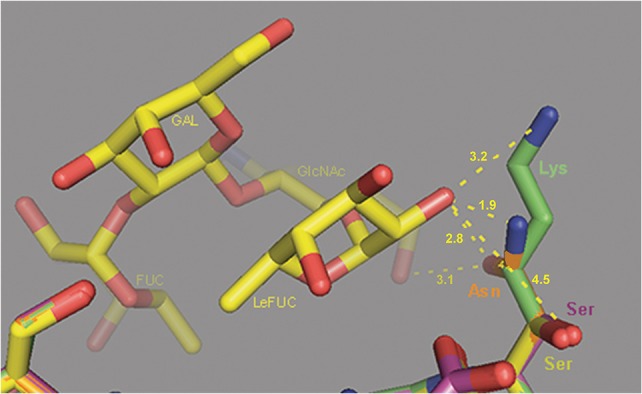
Noroviruses are the leading cause of acute gastroenteritis, and they can affect humans of all age groups. In immunocompromised patients, norovirus infections can develop into chronic diarrhea or show prolonged asymptomatic virus shedding. Chronic norovirus infections are frequently reported for solid organ transplant recipients, with rapid intrahost norovirus evolution seen. In this report, we describe a case of chronic norovirus infection in an immunocompromised patient who was followed up for over 5 years. The purpose of the study was to specify the norovirus evolution in a chronically infected immunocompromised host and identify possible selection sites in norovirus capsid protein. During the follow-up period, 25 sequential stool samples were collected and nine of them were selected to generate amplicons covering viral RNA-dependent RNA polymerase (RdRp) and viral capsid protein (VP1) genes. Amplicons were sequenced using next-generation sequencing. Single nucleotide polymorphisms were defined, which demonstrated a nearly 3-fold greater mutation rate in the VP1 genome region compared to the RdRp genome region (7.9 vs. 2.8 variable sites/100 nucleotides, respectively). This indicates that mutations in the virus genome were not accumulated randomly, but are rather the result of mutant selection during the infection cycle. Using ShoRAH software we were able to reconstruct haplotypes occurring in each of the nine selected samples. The deduced amino-acid haplotype sequences were aligned and the positions were analyzed for selective pressure using the Datamonkey program. Only 12 out of 25 positive selection sites were within the commonly described epitopes A, B, C, and D of the VP1 protein. New positive selection sites were determined that have not been described before and might reflect adaptation of the norovirus toward optimal histo-blood-group antigen binding, or modification of the norovirus antigenic properties. These data provide new insights into norovirus evolutionary dynamics and indicate new putative epitope "hot-spots" of modified and optimized norovirus-host interactions.
Keywords: HBGA binding; chronic infection; molecular evolution; norovirus; solid organ transplantation.
Figures
Similar articles
-
Long Term Norovirus Infection in a Patient with Severe Common Variable Immunodeficiency.Viruses. 2022 Aug 2;14(8):1708. doi: 10.3390/v14081708. Viruses. 2022. PMID: 36016330 Free PMC article.
-
Within-host evolution of virus variants during chronic infection with novel GII.P26-GII.26 norovirus.J Clin Virol. 2018 Nov;108:96-102. doi: 10.1016/j.jcv.2018.09.013. Epub 2018 Sep 19. J Clin Virol. 2018. PMID: 30268000
-
A next generation sequencing-based method to study the intra-host genetic diversity of norovirus in patients with acute and chronic infection.BMC Genomics. 2016 Jul 1;17:480. doi: 10.1186/s12864-016-2831-y. BMC Genomics. 2016. PMID: 27363999 Free PMC article.
-
Prolonged norovirus infection after pancreas transplantation: a case report and review of chronic norovirus.Transpl Infect Dis. 2016 Feb;18(1):98-104. doi: 10.1111/tid.12472. Epub 2015 Nov 25. Transpl Infect Dis. 2016. PMID: 26460906 Review.
-
The Antigenic Topology of Norovirus as Defined by B and T Cell Epitope Mapping: Implications for Universal Vaccines and Therapeutics.Viruses. 2019 May 10;11(5):432. doi: 10.3390/v11050432. Viruses. 2019. PMID: 31083353 Free PMC article. Review.
Cited by
-
Successfully Treated Norovirus- and Sapovirus-Associated Diarrhea in Three Renal Transplant Patients.Case Rep Infect Dis. 2018 Nov 12;2018:6846873. doi: 10.1155/2018/6846873. eCollection 2018. Case Rep Infect Dis. 2018. PMID: 30538873 Free PMC article.
-
Intra-Host Evolution of Norovirus GII.4 in a Chronic Infected Patient With Hematopoietic Stem Cell Transplantation.Front Microbiol. 2020 Mar 9;11:375. doi: 10.3389/fmicb.2020.00375. eCollection 2020. Front Microbiol. 2020. PMID: 32210947 Free PMC article.
-
Obesity-Related Microenvironment Promotes Emergence of Virulent Influenza Virus Strains.mBio. 2020 Mar 3;11(2):e03341-19. doi: 10.1128/mBio.03341-19. mBio. 2020. PMID: 32127459 Free PMC article.
-
Norovirus in health care and implications for the immunocompromised host.Curr Opin Infect Dis. 2019 Aug;32(4):348-355. doi: 10.1097/QCO.0000000000000557. Curr Opin Infect Dis. 2019. PMID: 31107251 Free PMC article.
-
Long Term Norovirus Infection in a Patient with Severe Common Variable Immunodeficiency.Viruses. 2022 Aug 2;14(8):1708. doi: 10.3390/v14081708. Viruses. 2022. PMID: 36016330 Free PMC article.
References
-
- Daniels G. (2002). ABO, H and lewis systems, in Human Blood Groups, ed Daniels G. (Oxford: Blackwell Science Ltd.), 11–95.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials