

Cooperative recruitment of Yan via a high-affinity ETS supersite organizes repression to confer specificity and robustness to cardiac cell fate specification
- PMID: 29535190
- PMCID: PMC5900712
- DOI: 10.1101/gad.307132.117
Cooperative recruitment of Yan via a high-affinity ETS supersite organizes repression to confer specificity and robustness to cardiac cell fate specification
Abstract
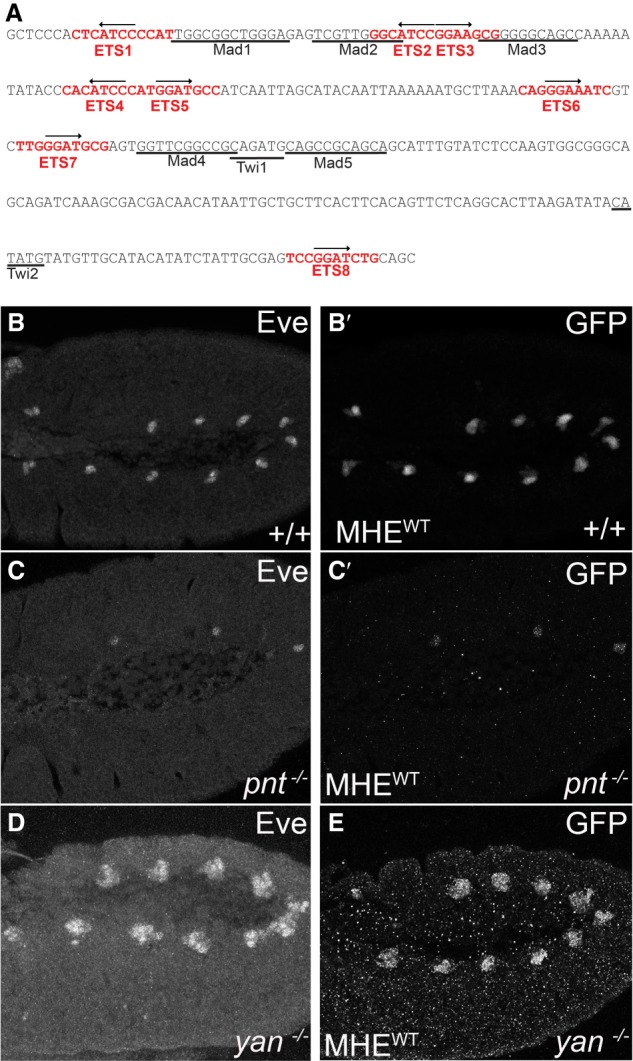
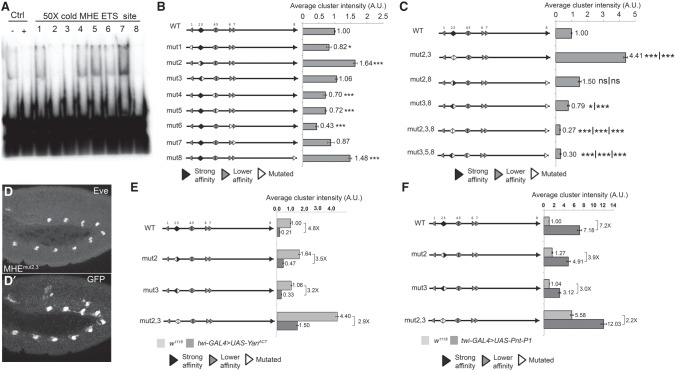
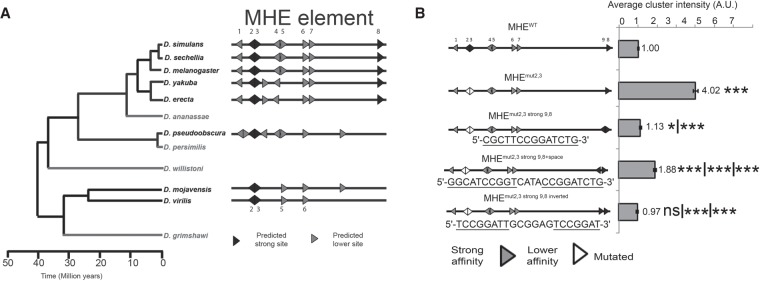
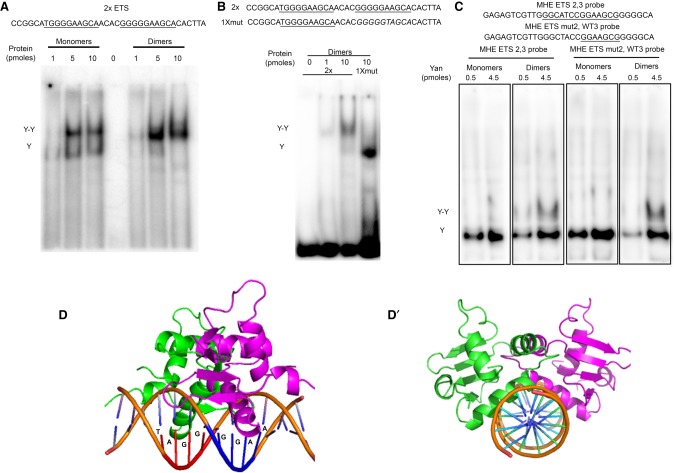
Cis-regulatory modules (CRMs) are defined by unique combinations of transcription factor-binding sites. Emerging evidence suggests that the number, affinity, and organization of sites play important roles in regulating enhancer output and, ultimately, gene expression. Here, we investigate how the cis-regulatory logic of a tissue-specific CRM responsible for even-skipped (eve) induction during cardiogenesis organizes the competing inputs of two E-twenty-six (ETS) members: the activator Pointed (Pnt) and the repressor Yan. Using a combination of reporter gene assays and CRISPR-Cas9 gene editing, we suggest that Yan and Pnt have distinct syntax preferences. Not only does Yan prefer high-affinity sites, but an overlapping pair of such sites is necessary and sufficient for Yan to tune Eve expression levels in newly specified cardioblasts and block ectopic Eve induction and cell fate specification in surrounding progenitors. Mechanistically, the efficient Yan recruitment promoted by this high-affinity ETS supersite not only biases Yan-Pnt competition at the specific CRM but also organizes Yan-repressive complexes in three dimensions across the eve locus. Taken together, our results uncover a novel mechanism by which differential interpretation of CRM syntax by a competing repressor-activator pair can confer both specificity and robustness to developmental transitions.
Keywords: Drosophila embryogenesis; ETS transcription factor; cis-regulatory syntax; even-skipped; heart development; receptor tyrosine kinase.
© 2018 Boisclair Lachance et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
Collaborative repressive action of the antagonistic ETS transcription factors Pointed and Yan fine-tunes gene expression to confer robustness in Drosophila.Development. 2018 Jul 2;145(13):dev165985. doi: 10.1242/dev.165985. Development. 2018. PMID: 29848501 Free PMC article.
-
Yan regulates Lozenge during Drosophila eye development.Dev Genes Evol. 2002 Jul;212(6):267-76. doi: 10.1007/s00427-002-0241-4. Epub 2002 Jun 13. Dev Genes Evol. 2002. PMID: 12111211
-
3D chromatin interactions organize Yan chromatin occupancy and repression at the even-skipped locus.Genes Dev. 2013 Nov 1;27(21):2293-8. doi: 10.1101/gad.225789.113. Genes Dev. 2013. PMID: 24186975 Free PMC article.
-
Proteins of the ETS family with transcriptional repressor activity.Oncogene. 2000 Dec 18;19(55):6524-32. doi: 10.1038/sj.onc.1204045. Oncogene. 2000. PMID: 11175368 Review.
-
Sequence and functional properties of Ets genes in the model organism Drosophila.Oncogene. 2000 Dec 18;19(55):6409-16. doi: 10.1038/sj.onc.1204033. Oncogene. 2000. PMID: 11175357 Review.
Cited by
-
Collaborative repressive action of the antagonistic ETS transcription factors Pointed and Yan fine-tunes gene expression to confer robustness in Drosophila.Development. 2018 Jul 2;145(13):dev165985. doi: 10.1242/dev.165985. Development. 2018. PMID: 29848501 Free PMC article.
-
Ratiometric sensing of Pnt and Yan transcription factor levels confers ultrasensitivity to photoreceptor fate transitions in Drosophila.Development. 2023 Apr 15;150(8):dev201467. doi: 10.1242/dev.201467. Epub 2023 Apr 24. Development. 2023. PMID: 36942737 Free PMC article.
-
Enhancer grammar in development, evolution, and disease: dependencies and interplay.Dev Cell. 2021 Mar 8;56(5):575-587. doi: 10.1016/j.devcel.2021.02.016. Dev Cell. 2021. PMID: 33689769 Free PMC article. Review.
-
Mutagenesis Sensitivity Mapping of Human Enhancers In Vivo.bioRxiv [Preprint]. 2024 Sep 8:2024.09.06.611737. doi: 10.1101/2024.09.06.611737. bioRxiv. 2024. PMID: 39282388 Free PMC article. Preprint.
-
ETS transcription factors regulate precise matrix metalloproteinase expression and follicle rupture in Drosophila.Development. 2024 Mar 1;151(5):dev202276. doi: 10.1242/dev.202276. Epub 2024 Feb 28. Development. 2024. PMID: 38345299
References
-
- Arnold K, Bordoli L, Kopp J, Schwede T. 2006. The SWISS-MODEL workspace: a Web-based environment for protein structure homology modelling. Bioinformatics 22: 195–201. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials