

Mechanism of inhibition of retromer transport by the bacterial effector RidL
- PMID: 29386389
- PMCID: PMC5816186
- DOI: 10.1073/pnas.1717383115
Mechanism of inhibition of retromer transport by the bacterial effector RidL
Abstract
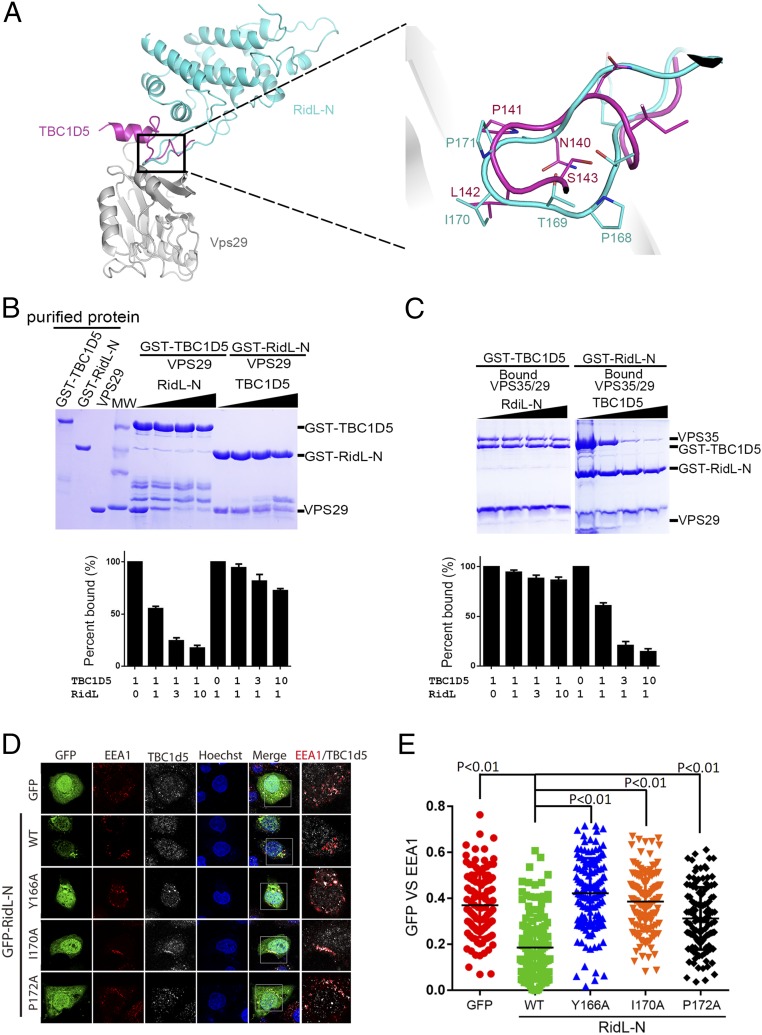
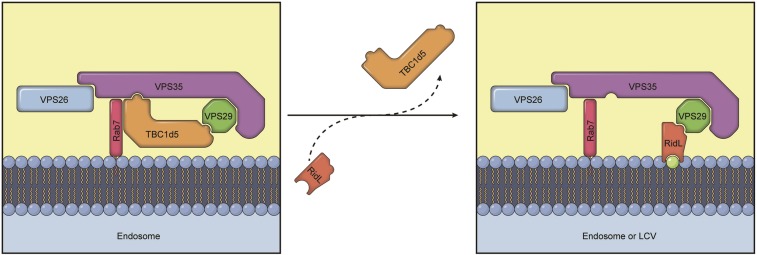
Retrograde vesicle trafficking pathways are responsible for returning membrane-associated components from endosomes to the Golgi apparatus and the endoplasmic reticulum (ER), and they are critical for maintaining organelle identity, lipid homeostasis, and many other cellular functions. The retrograde transport pathway has emerged as an important target for intravacuolar bacterial pathogens. The opportunistic pathogen Legionella pneumophila exploits both the secretory and recycling branches of the vesicle transport pathway for intracellular bacterial proliferation. Its Dot/Icm effector RidL inhibits the activity of the retromer by directly engaging retromer components. However, the mechanism underlying such inhibition remains unknown. Here we present the crystal structure of RidL in complex with VPS29, a subunit of the retromer. Our results demonstrate that RidL binds to a highly conserved hydrophobic pocket of VPS29. This interaction is critical for endosomal recruitment of RidL and for its inhibitory effects. RidL inhibits retromer activity by direct competition, in which it occupies the VPS29-binding site of the essential retromer regulator TBC1d5. The mechanism of retromer inhibition by RidL reveals a hotspot on VPS29 critical for recognition by its regulators that is also exploited by pathogens, and provides a structural basis for the development of small molecule inhibitors against the retromer.
Keywords: endosomal sorting; host–pathogen interaction; retromer; vesicular trafficking.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Structural insights into Legionella RidL-Vps29 retromer subunit interaction reveal displacement of the regulator TBC1D5.Nat Commun. 2017 Nov 16;8(1):1543. doi: 10.1038/s41467-017-01512-5. Nat Commun. 2017. PMID: 29146912 Free PMC article.
-
Molecular mechanism for the subversion of the retromer coat by the Legionella effector RidL.Proc Natl Acad Sci U S A. 2017 Dec 26;114(52):E11151-E11160. doi: 10.1073/pnas.1715361115. Epub 2017 Dec 11. Proc Natl Acad Sci U S A. 2017. PMID: 29229824 Free PMC article.
-
The Legionella effector RidL inhibits retrograde trafficking to promote intracellular replication.Cell Host Microbe. 2013 Jul 17;14(1):38-50. doi: 10.1016/j.chom.2013.06.001. Cell Host Microbe. 2013. PMID: 23870312
-
Formation of the Legionella Replicative Compartment at the Crossroads of Retrograde Trafficking.Front Cell Infect Microbiol. 2017 Nov 24;7:482. doi: 10.3389/fcimb.2017.00482. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 29226112 Free PMC article. Review.
-
Emerging Role of Retromer in Modulating Pathogen Growth.Trends Microbiol. 2018 Sep;26(9):769-780. doi: 10.1016/j.tim.2018.04.001. Epub 2018 Apr 24. Trends Microbiol. 2018. PMID: 29703496 Free PMC article. Review.
Cited by
-
Exploring the Diversity Within the Genus Francisella - An Integrated Pan-Genome and Genome-Mining Approach.Front Microbiol. 2020 Aug 11;11:1928. doi: 10.3389/fmicb.2020.01928. eCollection 2020. Front Microbiol. 2020. PMID: 32849479 Free PMC article.
-
Dictyostelium Dynamin Superfamily GTPases Implicated in Vesicle Trafficking and Host-Pathogen Interactions.Front Cell Dev Biol. 2021 Oct 13;9:731964. doi: 10.3389/fcell.2021.731964. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34746129 Free PMC article. Review.
-
Mechanism of cargo recognition by retromer-linked SNX-BAR proteins.PLoS Biol. 2020 Mar 9;18(3):e3000631. doi: 10.1371/journal.pbio.3000631. eCollection 2020 Mar. PLoS Biol. 2020. PMID: 32150533 Free PMC article.
-
Study of Legionella Effector Domains Revealed Novel and Prevalent Phosphatidylinositol 3-Phosphate Binding Domains.Infect Immun. 2019 May 21;87(6):e00153-19. doi: 10.1128/IAI.00153-19. Print 2019 Jun. Infect Immun. 2019. PMID: 30962397 Free PMC article.
-
FAM91A1-TBC1D23 complex structure reveals human genetic variations susceptible for PCH.Proc Natl Acad Sci U S A. 2023 Nov 7;120(45):e2309910120. doi: 10.1073/pnas.2309910120. Epub 2023 Oct 30. Proc Natl Acad Sci U S A. 2023. PMID: 37903274 Free PMC article.
References
-
- Lucas M, Hierro A. Retromer. Curr Biol. 2017;27:R687–R689. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
