

Nutritional Regulation of the Sae Two-Component System by CodY in Staphylococcus aureus
- PMID: 29378891
- PMCID: PMC5869476
- DOI: 10.1128/JB.00012-18
Nutritional Regulation of the Sae Two-Component System by CodY in Staphylococcus aureus
Abstract
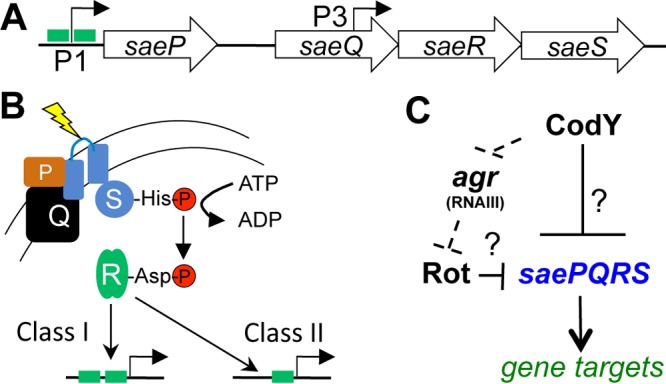
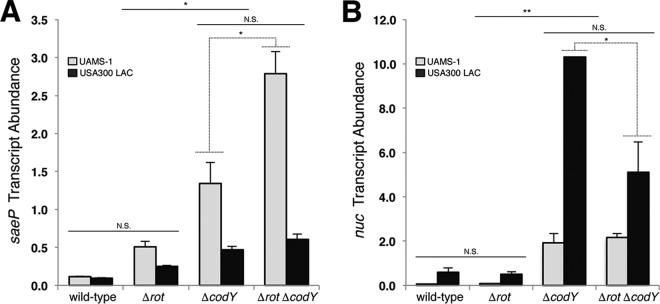
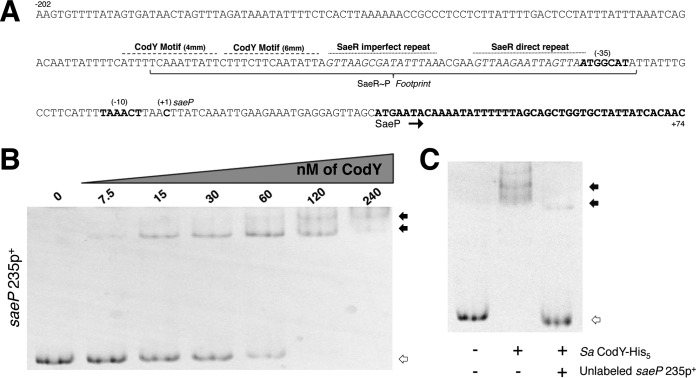
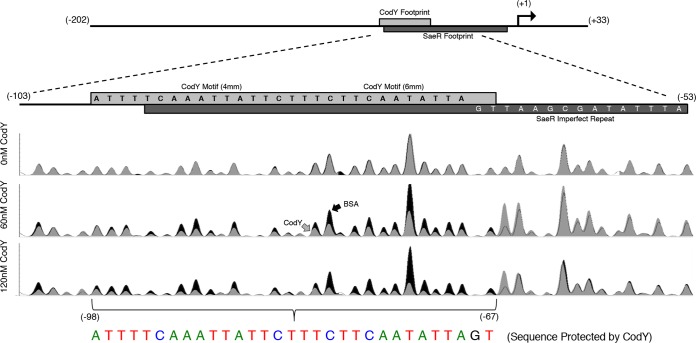
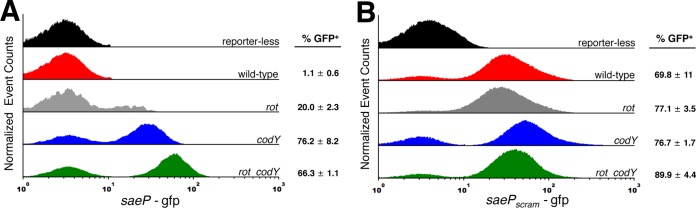
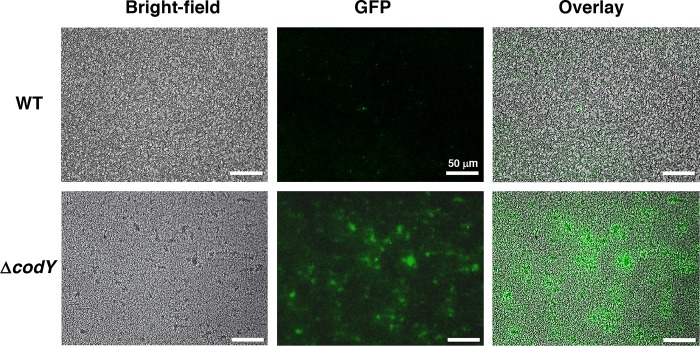
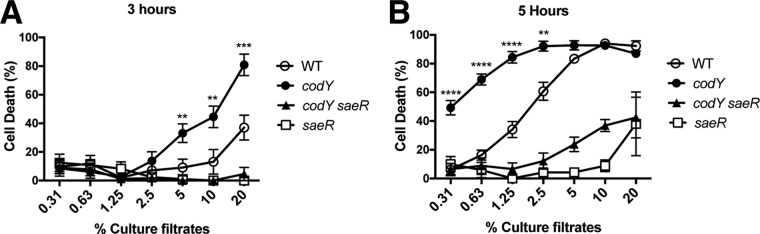
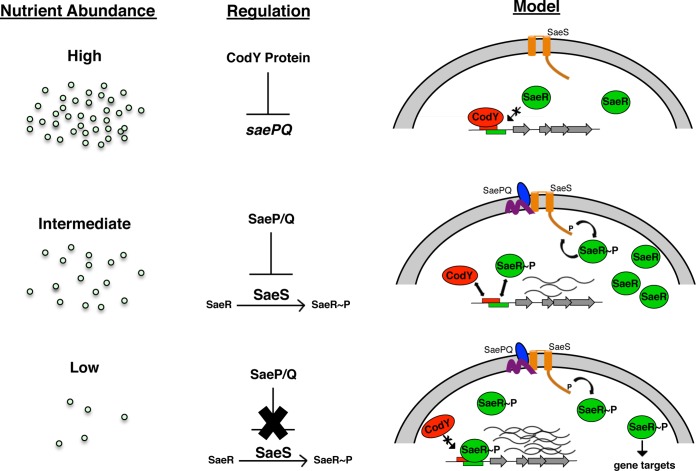
Staphylococcus aureus subverts innate defenses during infection in part by killing host immune cells to exacerbate disease. This human pathogen intercepts host cues and activates a transcriptional response via the S. aureus exoprotein expression (SaeR/SaeS [SaeR/S]) two-component system to secrete virulence factors critical for pathogenesis. We recently showed that the transcriptional repressor CodY adjusts nuclease (nuc) gene expression via SaeR/S, but the mechanism remained unknown. Here, we identified two CodY binding motifs upstream of the sae P1 promoter, which suggested direct regulation by this global regulator. We show that CodY shares a binding site with the positive activator SaeR and that alleviating direct CodY repression at this site is sufficient to abrogate stochastic expression, suggesting that CodY represses sae expression by blocking SaeR binding. Epistasis experiments support a model that CodY also controls sae indirectly through Agr and Rot-mediated repression of the sae P1 promoter. We also demonstrate that CodY repression of sae restrains production of secreted cytotoxins that kill human neutrophils. We conclude that CodY plays a previously unrecognized role in controlling virulence gene expression via SaeR/S and suggest a mechanism by which CodY acts as a master regulator of pathogenesis by tying nutrient availability to virulence gene expression.IMPORTANCE Bacterial mechanisms that mediate the switch from a commensal to pathogenic lifestyle are among the biggest unanswered questions in infectious disease research. Since the expression of most virulence genes is often correlated with nutrient depletion, this implies that virulence is a response to the lack of nourishment in host tissues and that pathogens like S. aureus produce virulence factors in order to gain access to nutrients in the host. Here, we show that specific nutrient depletion signals appear to be funneled to the SaeR/S system through the global regulator CodY. Our findings reveal a strategy by which S. aureus delays the production of immune evasion and immune-cell-killing proteins until key nutrients are depleted.
Keywords: CodY; Sae; Sae TCS; Staphylococcus aureus; branched-chain amino acids; gene regulation; two-component system; virulence.
Copyright © 2018 American Society for Microbiology.
Figures
Similar articles
-
Regulation of the Sae Two-Component System by Branched-Chain Fatty Acids in Staphylococcus aureus.mBio. 2022 Oct 26;13(5):e0147222. doi: 10.1128/mbio.01472-22. Epub 2022 Sep 22. mBio. 2022. PMID: 36135382 Free PMC article.
-
CodY controls the SaeR/S two-component system by modulating branched-chain fatty acid synthesis in Staphylococcus aureus.J Bacteriol. 2024 Nov 21;206(11):e0019124. doi: 10.1128/jb.00191-24. Epub 2024 Oct 9. J Bacteriol. 2024. PMID: 39382300
-
A spectrum of CodY activities drives metabolic reorganization and virulence gene expression in Staphylococcus aureus.Mol Microbiol. 2016 Aug;101(3):495-514. doi: 10.1111/mmi.13404. Epub 2016 Jun 10. Mol Microbiol. 2016. PMID: 27116338 Free PMC article.
-
The CodY pleiotropic repressor controls virulence in gram-positive pathogens.FEMS Immunol Med Microbiol. 2011 Jul;62(2):123-39. doi: 10.1111/j.1574-695X.2011.00812.x. Epub 2011 May 27. FEMS Immunol Med Microbiol. 2011. PMID: 21539625 Review.
-
CodY, a master integrator of metabolism and virulence in Gram-positive bacteria.Curr Genet. 2017 Jun;63(3):417-425. doi: 10.1007/s00294-016-0656-5. Epub 2016 Oct 15. Curr Genet. 2017. PMID: 27744611 Review.
Cited by
-
Repression of branched-chain amino acid synthesis in Staphylococcus aureus is mediated by isoleucine via CodY, and by a leucine-rich attenuator peptide.PLoS Genet. 2018 Jan 22;14(1):e1007159. doi: 10.1371/journal.pgen.1007159. eCollection 2018 Jan. PLoS Genet. 2018. PMID: 29357354 Free PMC article.
-
Quorum Sensing and Toxin Production in Staphylococcus aureus Osteomyelitis: Pathogenesis and Paradox.Toxins (Basel). 2020 Aug 12;12(8):516. doi: 10.3390/toxins12080516. Toxins (Basel). 2020. PMID: 32806558 Free PMC article. Review.
-
The general stress response of Staphylococcus aureus promotes tolerance of antibiotics and survival in whole human blood.Microbiology (Reading). 2020 Nov;166(11):1088-1094. doi: 10.1099/mic.0.000983. Epub 2020 Oct 23. Microbiology (Reading). 2020. PMID: 33095698 Free PMC article.
-
Two-Component Systems of S. aureus: Signaling and Sensing Mechanisms.Genes (Basel). 2021 Dec 23;13(1):34. doi: 10.3390/genes13010034. Genes (Basel). 2021. PMID: 35052374 Free PMC article. Review.
-
Regulation of the Sae Two-Component System by Branched-Chain Fatty Acids in Staphylococcus aureus.mBio. 2022 Oct 26;13(5):e0147222. doi: 10.1128/mbio.01472-22. Epub 2022 Sep 22. mBio. 2022. PMID: 36135382 Free PMC article.
References
-
- Gorwitz RJ, Kruszon-Moran D, McAllister SK, McQuillan G, McDougal LK, Fosheim GE, Jensen BJ, Killgore G, Tenover FC, Kuehnert MJ. 2008. Changes in the prevalence of nasal colonization with Staphylococcus aureus in the United States, 2001–2004. J Infect Dis 197:1226–1234. doi:10.1086/533494. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
