

New functions of Semaphorin 3E and its receptor PlexinD1 during developing and adult hippocampal formation
- PMID: 29358640
- PMCID: PMC5777998
- DOI: 10.1038/s41598-018-19794-0
New functions of Semaphorin 3E and its receptor PlexinD1 during developing and adult hippocampal formation
Abstract
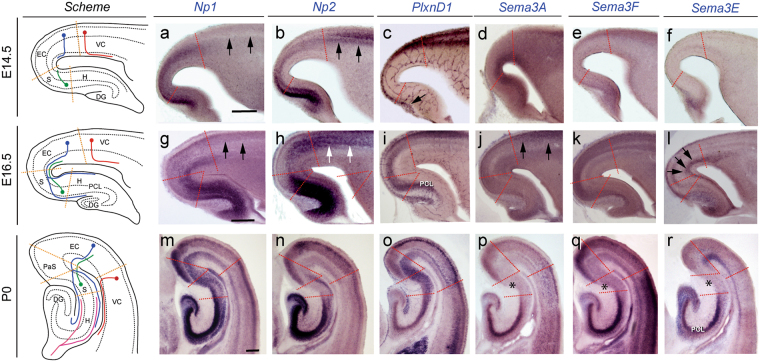
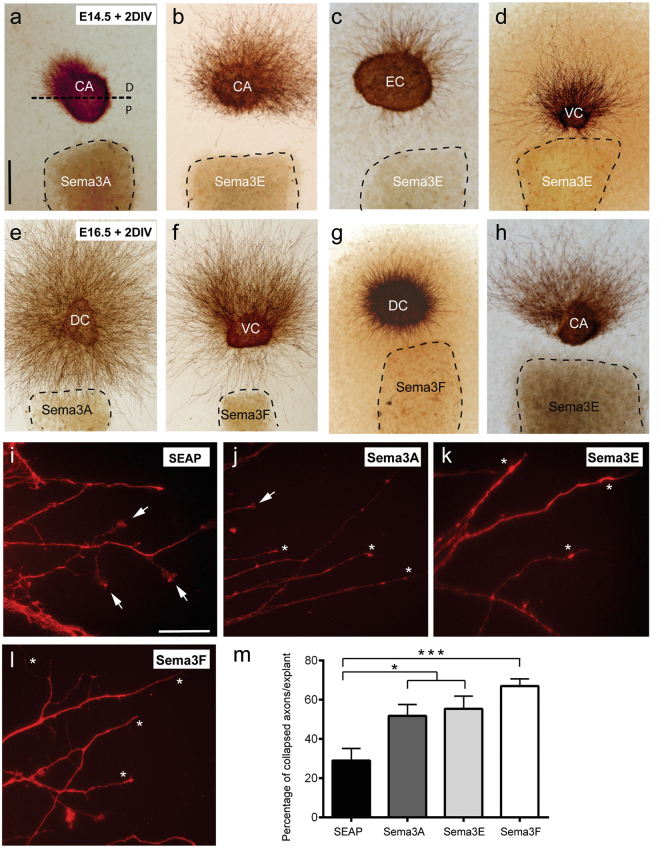
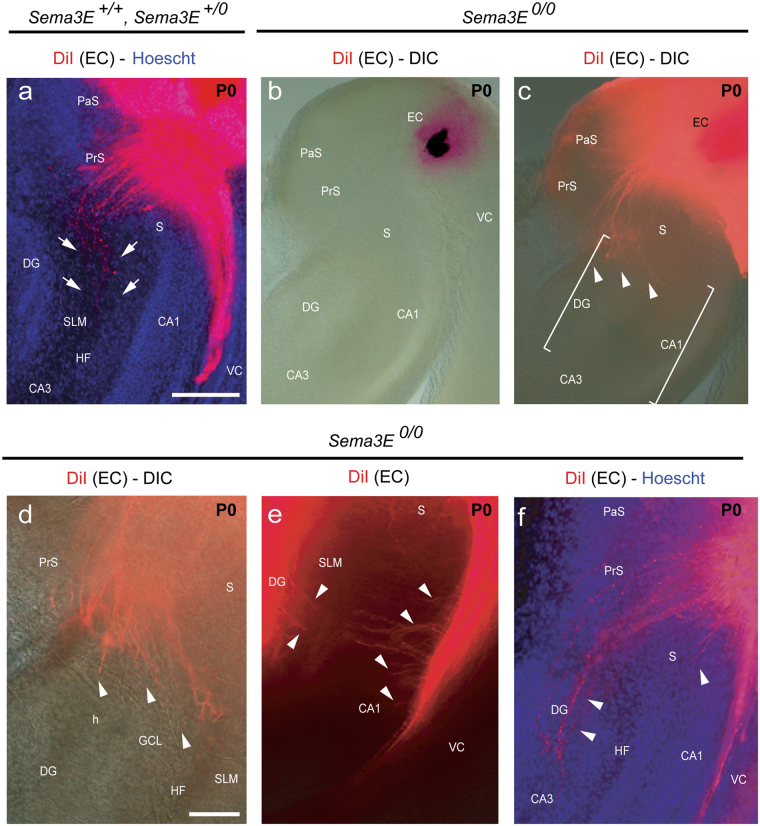
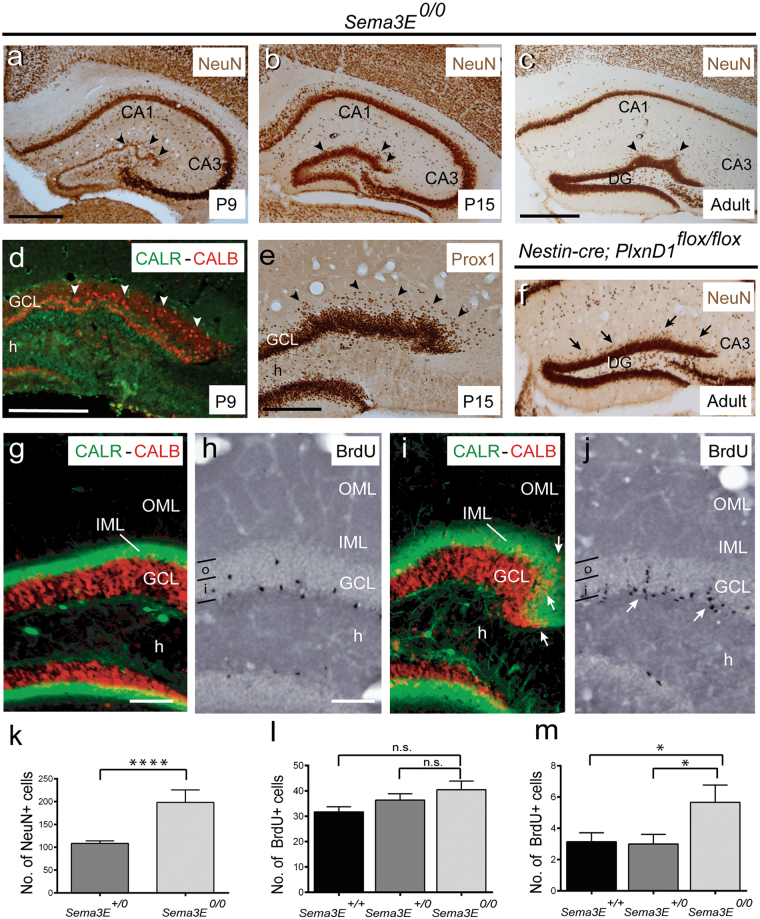
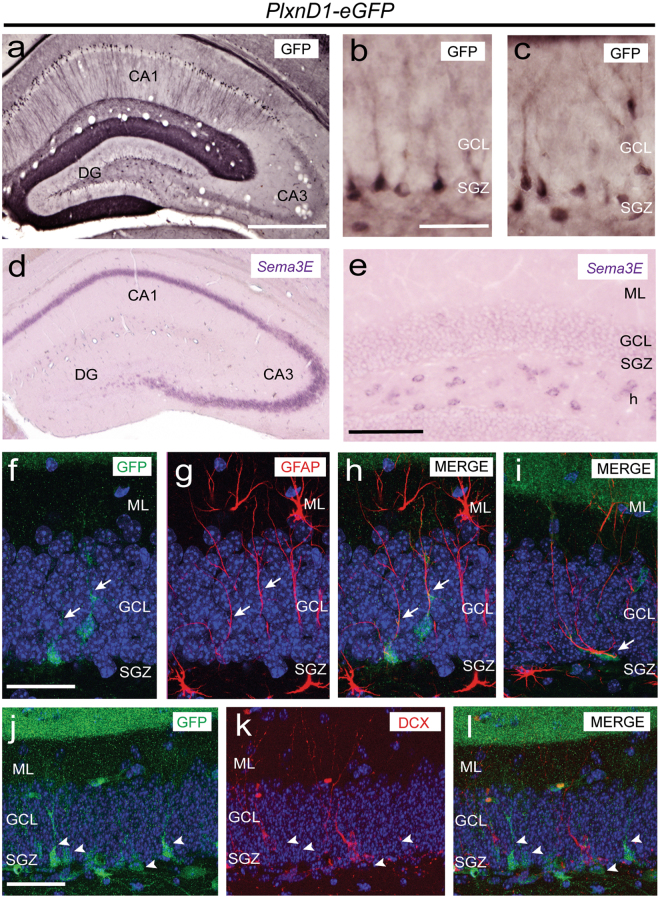
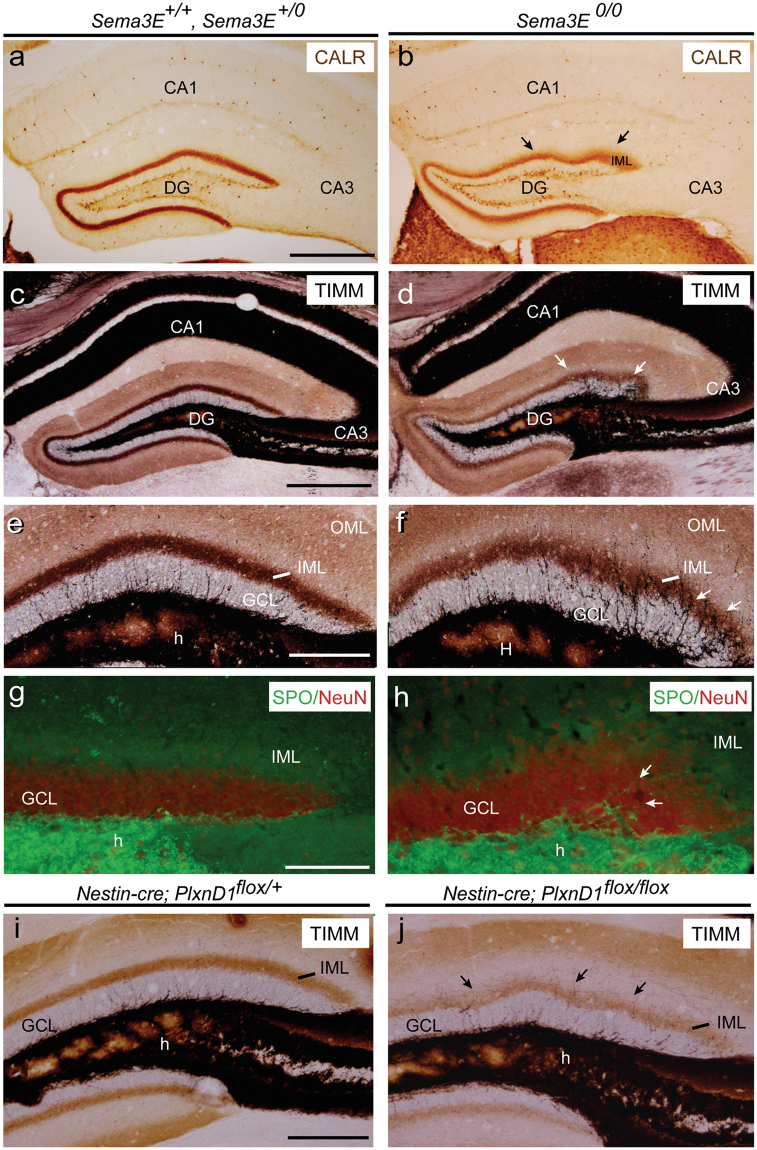
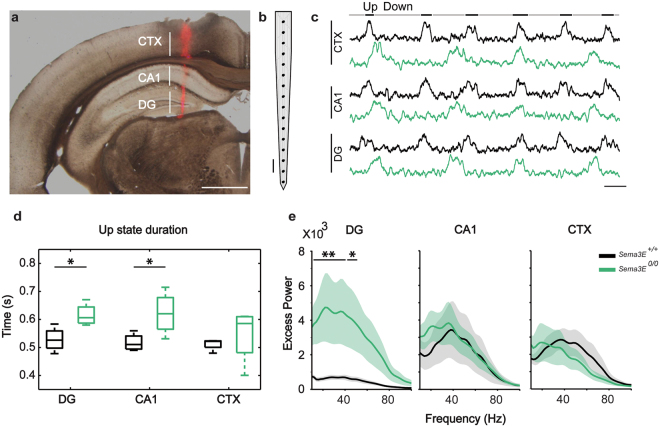
The development and maturation of cortical circuits relies on the coordinated actions of long and short range axonal guidance cues. In this regard, the class 3 semaphorins and their receptors have been seen to be involved in the development and maturation of the hippocampal connections. However, although the role of most of their family members have been described, very few data about the participation of Semaphorin 3E (Sema3E) and its receptor PlexinD1 during the development and maturation of the entorhino-hippocampal (EH) connection are available. In the present study, we focused on determining their roles both during development and in adulthood. We determined a relevant role for Sema3E/PlexinD1 in the layer-specific development of the EH connection. Indeed, mice lacking Sema3E/PlexinD1 signalling showed aberrant layering of entorhinal axons in the hippocampus during embryonic and perinatal stages. In addition, absence of Sema3E/PlexinD1 signalling results in further changes in postnatal and adult hippocampal formation, such as numerous misrouted ectopic mossy fibers. More relevantly, we describe how subgranular cells express PlexinD1 and how the absence of Sema3E induces a dysregulation of the proliferation of dentate gyrus progenitors leading to the presence of ectopic cells in the molecular layer. Lastly, Sema3E mutant mice displayed increased network excitability both in the dentate gyrus and the hippocampus proper.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Sema3E/PlexinD1 regulates the migration of hem-derived Cajal-Retzius cells in developing cerebral cortex.Nat Commun. 2014 Jun 27;5:4265. doi: 10.1038/ncomms5265. Nat Commun. 2014. PMID: 24969029
-
Age-dependent effects of secreted Semaphorins 3A, 3F, and 3E on developing hippocampal axons: in vitro effects and phenotype of Semaphorin 3A (-/-) mice.Mol Cell Neurosci. 2001 Jul;18(1):26-43. doi: 10.1006/mcne.2001.0999. Mol Cell Neurosci. 2001. PMID: 11461151
-
Gating of Sema3E/PlexinD1 signaling by neuropilin-1 switches axonal repulsion to attraction during brain development.Neuron. 2007 Dec 6;56(5):807-22. doi: 10.1016/j.neuron.2007.10.019. Neuron. 2007. PMID: 18054858 Free PMC article.
-
New molecules for hippocampal development.Trends Neurosci. 2001 Feb;24(2):107-13. doi: 10.1016/s0166-2236(00)01717-3. Trends Neurosci. 2001. PMID: 11164941 Review.
-
Sema-3E/PlexinD1 axis modulates dendritic cell phenotypes and functions: Current status and future implications.Hum Immunol. 2024 Jul;85(4):110815. doi: 10.1016/j.humimm.2024.110815. Epub 2024 May 20. Hum Immunol. 2024. PMID: 38772051 Review.
Cited by
-
Repulsive Sema3E-Plexin-D1 signaling coordinates both axonal extension and steering via activating an autoregulatory factor, Mtss1.Elife. 2024 Mar 25;13:e96891. doi: 10.7554/eLife.96891. Elife. 2024. PMID: 38526535 Free PMC article.
-
Involvement of Mechanical Cues in the Migration of Cajal-Retzius Cells in the Marginal Zone During Neocortical Development.Front Cell Dev Biol. 2022 May 16;10:886110. doi: 10.3389/fcell.2022.886110. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35652101 Free PMC article.
-
Roles and Mechanisms of Axon-Guidance Molecules in Alzheimer's Disease.Mol Neurobiol. 2021 Jul;58(7):3290-3307. doi: 10.1007/s12035-021-02311-2. Epub 2021 Mar 5. Mol Neurobiol. 2021. PMID: 33675023 Review.
-
NK/DC crosstalk-modulating antitumor activity via Sema3E/PlexinD1 axis for enhanced cancer immunotherapy.Immunol Res. 2024 Dec;72(6):1217-1228. doi: 10.1007/s12026-024-09536-y. Epub 2024 Sep 5. Immunol Res. 2024. PMID: 39235526 Review.
-
Semaphorin4A-Plexin D1 Axis Induces Th2 and Th17 While Represses Th1 Skewing in an Autocrine Manner.Int J Mol Sci. 2020 Sep 22;21(18):6965. doi: 10.3390/ijms21186965. Int J Mol Sci. 2020. PMID: 32971928 Free PMC article.
References
-
- Taupin, P. The hippocampus: neurotransmission and plasticity in the nervous system (Nova Biomedical Books, 2007).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
