

Leishmania proteophosphoglycans regurgitated from infected sand flies accelerate dermal wound repair and exacerbate leishmaniasis via insulin-like growth factor 1-dependent signalling
- PMID: 29352310
- PMCID: PMC5792026
- DOI: 10.1371/journal.ppat.1006794
Leishmania proteophosphoglycans regurgitated from infected sand flies accelerate dermal wound repair and exacerbate leishmaniasis via insulin-like growth factor 1-dependent signalling
Abstract
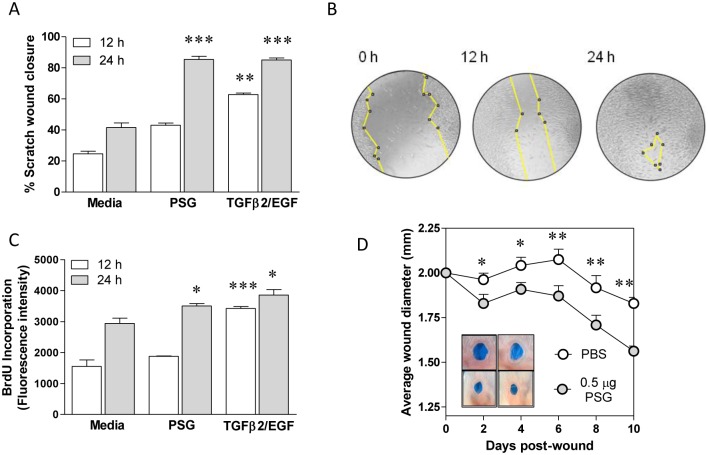
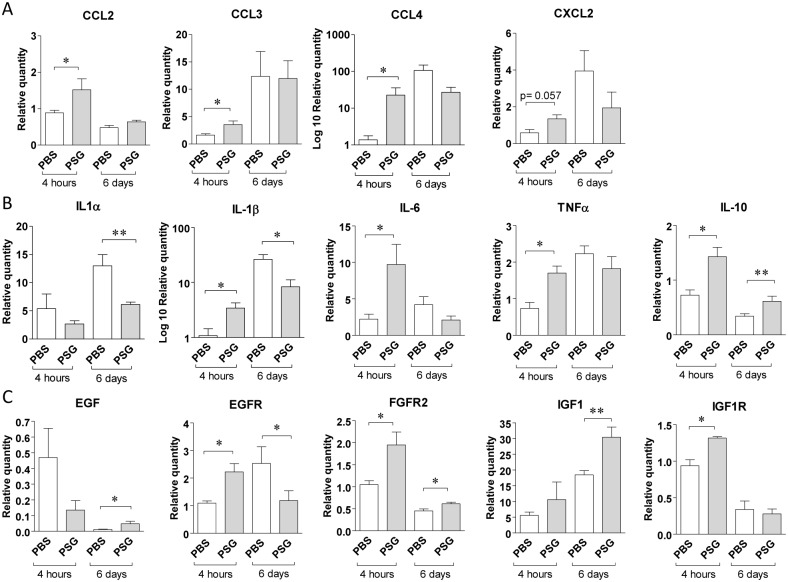
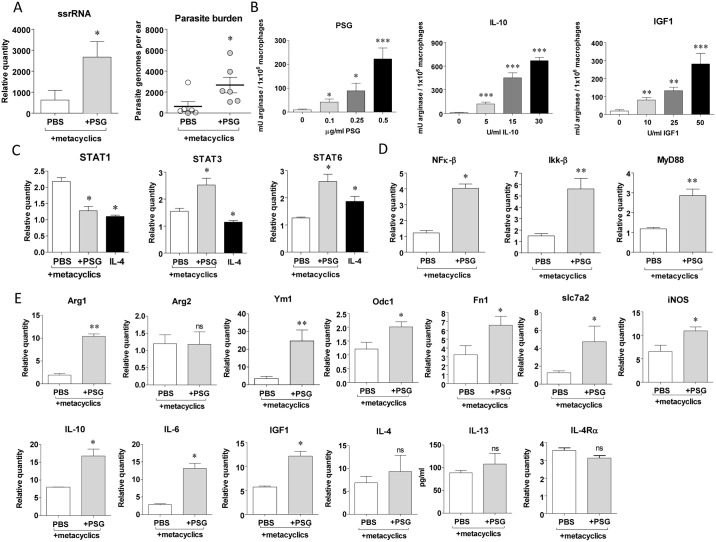
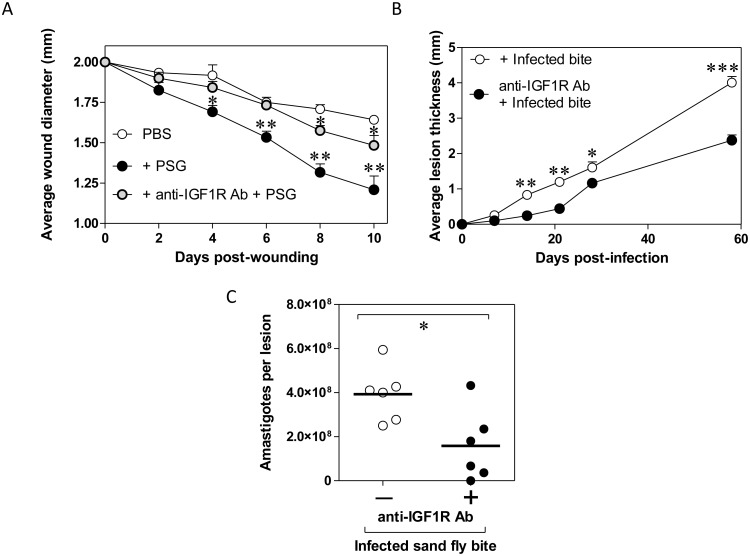
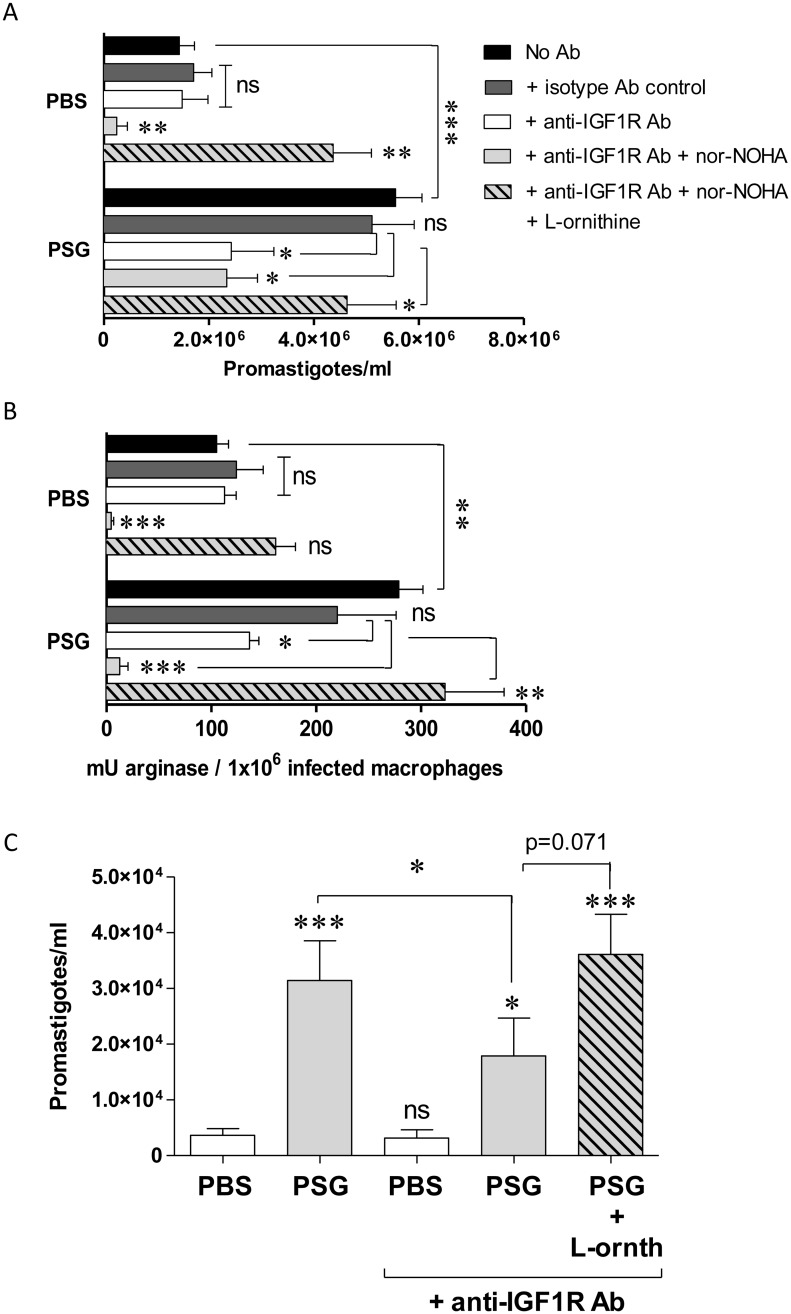
Leishmania parasites are transmitted to vertebrate hosts by female phlebotomine sand flies as they bloodfeed by lacerating the upper capillaries of the dermis with their barbed mouthparts. In the sand fly midgut secreted proteophosphoglycans from Leishmania form a biological plug known as the promastigote secretory gel (PSG), which blocks the gut and facilitates the regurgitation of infective parasites. The interaction between the wound created by the sand fly bite and PSG is not known. Here we nanoinjected a sand fly egested dose of PSG into BALB/c mouse skin that lead to the differential expression of 7,907 transcripts. These transcripts were transiently up-regulated during the first 6 hours post-wound and enriched for pathways involved in inflammation, cell proliferation, fibrosis, epithelial cell differentiation and wound remodelling. We found that PSG significantly accelerated wound healing in vitro and in mice; which was associated with an early up-regulation of transcripts involved in inflammation (IL-1β, IL-6, IL-10, TNFα) and inflammatory cell recruitment (CCL2, CCL3, CCL4, CXCL2), followed 6 days later by enhanced expression of transcripts associated with epithelial cell proliferation, fibroplasia and fibrosis (FGFR2, EGF, EGFR, IGF1). Dermal expression of IGF1 was enhanced following an infected sand fly bite and was acutely responsive to the deposition of PSG but not the inoculation of parasites or sand fly saliva. Antibody blockade of IGF1 ablated the gel's ability to promote wound closure in mouse ears and significantly reduced the virulence of Leishmania mexicana infection delivered by an individual sand fly bite. Dermal macrophages recruited to air-pouches on the backs of mice revealed that IGF1 was pivotal to the PSG's ability to promote macrophage alternative activation and Leishmania infection. Our data demonstrate that through the regurgitation of PSG Leishmania exploit the wound healing response of the host to the vector bite by promoting the action of IGF1 to drive the alternative activation of macrophages.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



Similar articles
-
Promastigote secretory gel from natural and unnatural sand fly vectors exacerbate Leishmania major and Leishmania tropica cutaneous leishmaniasis in mice.Parasitology. 2019 Dec;146(14):1796-1802. doi: 10.1017/S0031182019001069. Epub 2019 Aug 29. Parasitology. 2019. PMID: 31452467 Free PMC article.
-
Proteophosophoglycans regurgitated by Leishmania-infected sand flies target the L-arginine metabolism of host macrophages to promote parasite survival.PLoS Pathog. 2009 Aug;5(8):e1000555. doi: 10.1371/journal.ppat.1000555. Epub 2009 Aug 21. PLoS Pathog. 2009. PMID: 19696894 Free PMC article.
-
Leishmania infantum proteophosphoglycans regurgitated by the bite of its natural sand fly vector, Lutzomyia longipalpis, promote parasite establishment in mouse skin and skin-distant tissues.Microbes Infect. 2010 Oct;12(11):875-9. doi: 10.1016/j.micinf.2010.05.014. Epub 2010 Jun 16. Microbes Infect. 2010. PMID: 20561596
-
Transmission of Leishmania metacyclic promastigotes by phlebotomine sand flies.Int J Parasitol. 2007 Aug;37(10):1097-106. doi: 10.1016/j.ijpara.2007.04.003. Epub 2007 Apr 18. Int J Parasitol. 2007. PMID: 17517415 Free PMC article. Review.
-
Gels and cells: the Leishmania biofilm as a space and place for parasite transmission.Trends Parasitol. 2024 Oct;40(10):876-885. doi: 10.1016/j.pt.2024.08.001. Epub 2024 Aug 31. Trends Parasitol. 2024. PMID: 39218719 Review.
Cited by
-
Omics Approaches in Drug Development against Leishmaniasis: Current Scenario and Future Prospects.Pathogens. 2022 Dec 26;12(1):39. doi: 10.3390/pathogens12010039. Pathogens. 2022. PMID: 36678387 Free PMC article. Review.
-
Macrophage Polarization in Leishmaniasis: Broadening Horizons.Front Immunol. 2018 Oct 31;9:2529. doi: 10.3389/fimmu.2018.02529. eCollection 2018. Front Immunol. 2018. PMID: 30429856 Free PMC article. Review.
-
The First Non-LRV RNA Virus in Leishmania.Viruses. 2020 Feb 2;12(2):168. doi: 10.3390/v12020168. Viruses. 2020. PMID: 32024293 Free PMC article.
-
Promastigote secretory gel from natural and unnatural sand fly vectors exacerbate Leishmania major and Leishmania tropica cutaneous leishmaniasis in mice.Parasitology. 2019 Dec;146(14):1796-1802. doi: 10.1017/S0031182019001069. Epub 2019 Aug 29. Parasitology. 2019. PMID: 31452467 Free PMC article.
-
Collagen-Containing Fish Sidestream-Derived Protein Hydrolysates Support Skin Repair via Chemokine Induction.Mar Drugs. 2021 Jul 15;19(7):396. doi: 10.3390/md19070396. Mar Drugs. 2021. PMID: 34356821 Free PMC article.
References
-
- Sacks D, Kamhawi S (2001) Molecular aspects of parasite-vector and vector-host interactions in leishmaniasis. Annu Rev Microbiol 55: 453–483. doi: 10.1146/annurev.micro.55.1.453 - DOI - PubMed
-
- Lestinova T, Rohousova I, Sima M, de Oliveira CI, Volf P (2017) Insights into the sand fly saliva: Blood-feeding and immune interactions between sand flies, hosts, and Leishmania. PLoS Negl Trop Dis. 11: e0005600 doi: 10.1371/journal.pntd.0005600 - DOI - PMC - PubMed
-
- Titus RG, Ribeiro JM (1988) Salivary gland lysates from the sand fly Lutzomyia longipalpis enhanced Leishmania infectivity. Science 239: 1306–1308. - PubMed
-
- Kamhawi S, Belkaid Y, Modi G, Rowton E, Sacks DL (2000) Protection against cutaneous leishmaniasis resulting from bites of uninfected sand flies. Science 290: 1351–1354. - PubMed
-
- Oliveira F, Lawyer PG, Kamhawi S, Valenzuela JG (2008) Immunity to distinct sand fly salivary proteins primes the anti-Leishmania immune response toward protection or exacerbation of disease. PLoS Negl Trop Dis 2: e226 doi: 10.1371/journal.pntd.0000226 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
