

MitoNEET in Perivascular Adipose Tissue Blunts Atherosclerosis under Mild Cold Condition in Mice
- PMID: 29311966
- PMCID: PMC5742148
- DOI: 10.3389/fphys.2017.01032
MitoNEET in Perivascular Adipose Tissue Blunts Atherosclerosis under Mild Cold Condition in Mice
Abstract
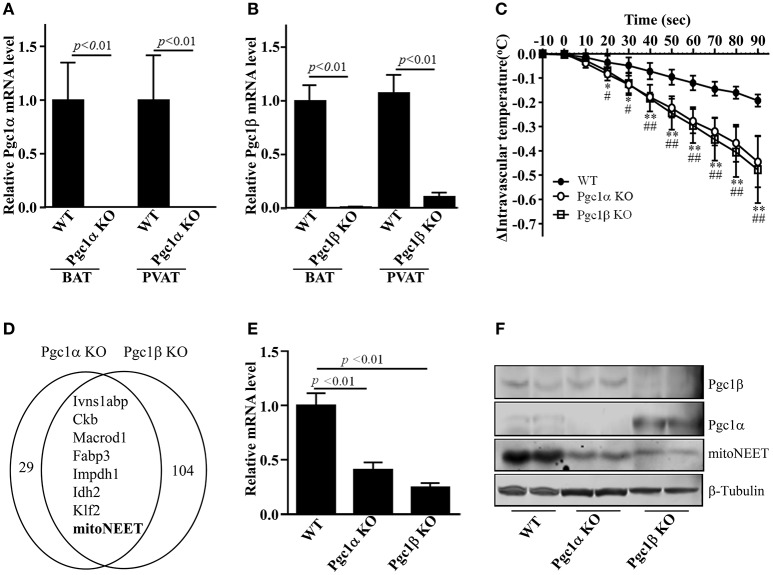
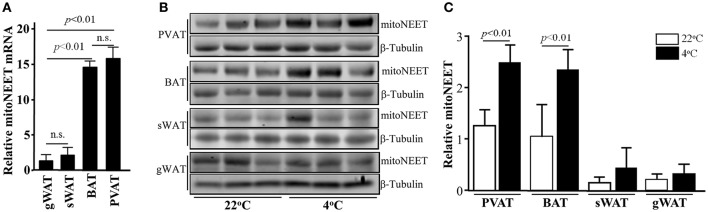
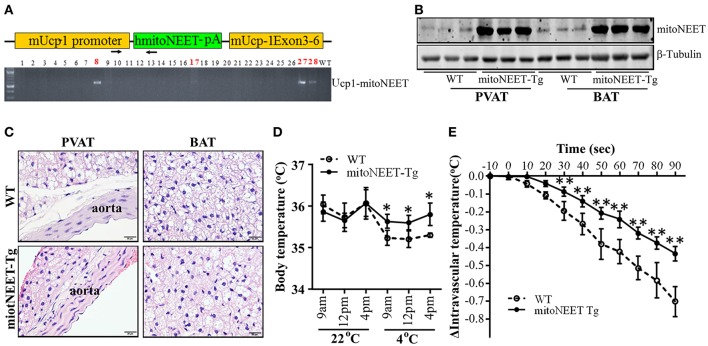
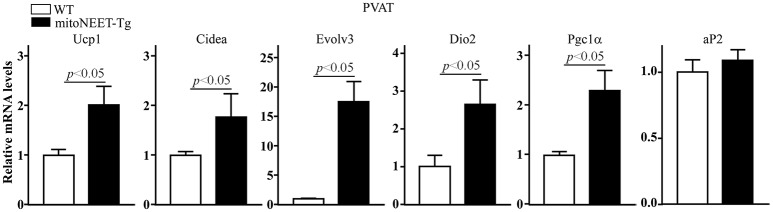
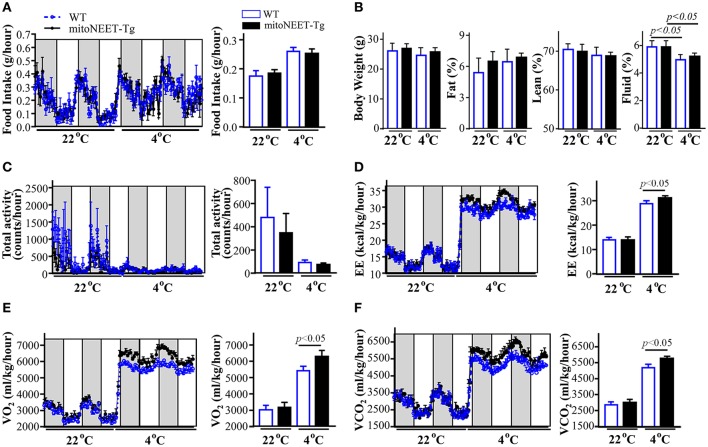
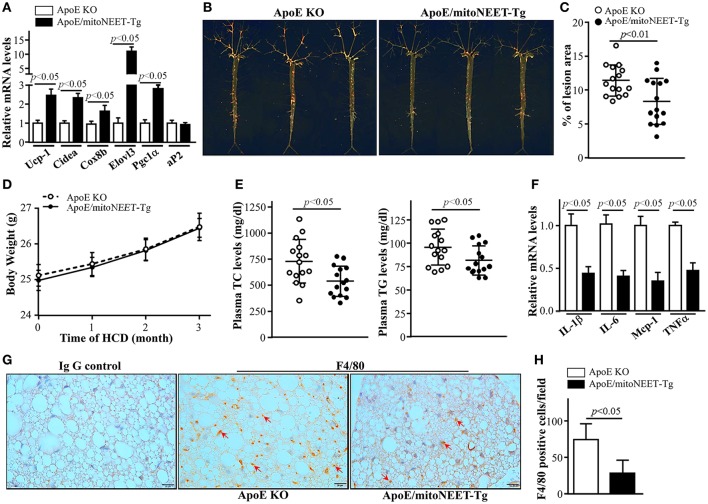
Background: Perivascular adipose tissue (PVAT), which surrounds most vessels, is de facto a distinct functional vascular layer actively contributing to vascular function and dysfunction. PVAT contributes to aortic remodeling by producing and releasing a large number of undetermined or less characterized factors that could target endothelial cells and vascular smooth muscle cells, and herein contribute to the maintenance of vessel homeostasis. Loss of PVAT in mice enhances atherosclerosis, but a causal relationship between PVAT and atherosclerosis and the possible underlying mechanisms remain to be addressed. The CDGSH iron sulfur domain 1 protein (referred to as mitoNEET), a mitochondrial outer membrane protein, regulates oxidative capacity and adipose tissue browning. The roles of mitoNEET in PVAT, especially in the development of atherosclerosis, are unknown. Methods: The brown adipocyte-specific mitoNEET transgenic mice were subjected to cold environmental stimulus. The metabolic rates and PVAT-dependent thermogenesis were investigated. Additionally, the brown adipocyte-specific mitoNEET transgenic mice were cross-bred with ApoE knockout mice. The ensuing mice were subsequently subjected to cold environmental stimulus and high cholesterol diet challenge for 3 months. The development of atherosclerosis was investigated. Results: Our data show that mitoNEET mRNA was downregulated in PVAT of both peroxisome proliferator-activated receptor gamma coactivator 1-alpha (Pgc1α)- and beta (Pgc1β)-knockout mice which are sensitive to cold. MitoNEET expression was higher in PVAT of wild type mice and increased upon cold stimulus. Transgenic mice with overexpression of mitoNEET in PVAT were cold resistant, and showed increased expression of thermogenic genes. ApoE knockout mice with mitoNEET overexpression in PVAT showed significant downregulation of inflammatory genes and showed reduced atherosclerosis development upon high fat diet feeding when kept in a 16°C environment. Conclusion: mitoNEET in PVAT is associated with PVAT-dependent thermogenesis and prevents atherosclerosis development. The results of this study provide new insights on PVAT and mitoNEET biology and atherosclerosis in cardiovascular diseases.
Keywords: Cisd1; atherosclerosis; mitoNEET; mitochondria; perivascular adipose tissue.
Figures
Similar articles
-
Roles of Perivascular Adipose Tissue in Hypertension and Atherosclerosis.Antioxid Redox Signal. 2021 Mar 20;34(9):736-749. doi: 10.1089/ars.2020.8103. Epub 2020 Jun 2. Antioxid Redox Signal. 2021. PMID: 32390459 Free PMC article. Review.
-
Brown Adipocyte-Specific PPARγ (Peroxisome Proliferator-Activated Receptor γ) Deletion Impairs Perivascular Adipose Tissue Development and Enhances Atherosclerosis in Mice.Arterioscler Thromb Vasc Biol. 2018 Aug;38(8):1738-1747. doi: 10.1161/ATVBAHA.118.311367. Arterioscler Thromb Vasc Biol. 2018. PMID: 29954752 Free PMC article.
-
MitoNEET in Perivascular Adipose Tissue Prevents Arterial Stiffness in Aging Mice.Cardiovasc Drugs Ther. 2018 Oct;32(5):531-539. doi: 10.1007/s10557-018-6809-7. Cardiovasc Drugs Ther. 2018. PMID: 30022354
-
Loss of perivascular adipose tissue on peroxisome proliferator-activated receptor-γ deletion in smooth muscle cells impairs intravascular thermoregulation and enhances atherosclerosis.Circulation. 2012 Aug 28;126(9):1067-78. doi: 10.1161/CIRCULATIONAHA.112.104489. Epub 2012 Aug 1. Circulation. 2012. PMID: 22855570 Free PMC article.
-
Perivascular adipose tissue (PVAT) in atherosclerosis: a double-edged sword.Cardiovasc Diabetol. 2018 Oct 10;17(1):134. doi: 10.1186/s12933-018-0777-x. Cardiovasc Diabetol. 2018. PMID: 30305178 Free PMC article. Review.
Cited by
-
Inhibition of a Novel CLK1-THRAP3-PPARγ Axis Improves Insulin Sensitivity.Front Physiol. 2021 Aug 30;12:699578. doi: 10.3389/fphys.2021.699578. eCollection 2021. Front Physiol. 2021. PMID: 34526909 Free PMC article.
-
Pathological Conversion of Mouse Perivascular Adipose Tissue by Notch Activation.Arterioscler Thromb Vasc Biol. 2020 Sep;40(9):2227-2243. doi: 10.1161/ATVBAHA.120.314731. Epub 2020 Jul 9. Arterioscler Thromb Vasc Biol. 2020. PMID: 32640901 Free PMC article.
-
Roles of Perivascular Adipose Tissue in Hypertension and Atherosclerosis.Antioxid Redox Signal. 2021 Mar 20;34(9):736-749. doi: 10.1089/ars.2020.8103. Epub 2020 Jun 2. Antioxid Redox Signal. 2021. PMID: 32390459 Free PMC article. Review.
-
Perivascular Adipose Tissue as a Target for Antioxidant Therapy for Cardiovascular Complications.Antioxidants (Basel). 2020 Jul 2;9(7):574. doi: 10.3390/antiox9070574. Antioxidants (Basel). 2020. PMID: 32630640 Free PMC article. Review.
-
Brown Adipocyte-Specific PPARγ (Peroxisome Proliferator-Activated Receptor γ) Deletion Impairs Perivascular Adipose Tissue Development and Enhances Atherosclerosis in Mice.Arterioscler Thromb Vasc Biol. 2018 Aug;38(8):1738-1747. doi: 10.1161/ATVBAHA.118.311367. Arterioscler Thromb Vasc Biol. 2018. PMID: 29954752 Free PMC article.
References
-
- Balogh L., Donhoffer S., Mestyan G., Pap T., Toth I. (1952). The effect of environmental temperature on the O2-consumption and body temperature of rats under the acute action of some drugs affecting energy exchange and body temperature. Acta Physiol. Acad. Sci. Hung. 3, 367–375. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous