

Genomic, Transcriptomic, and Phenotypic Analyses of Neisseria meningitidis Isolates from Disease Patients and Their Household Contacts
- PMID: 29152586
- PMCID: PMC5686521
- DOI: 10.1128/mSystems.00127-17
Genomic, Transcriptomic, and Phenotypic Analyses of Neisseria meningitidis Isolates from Disease Patients and Their Household Contacts
Abstract
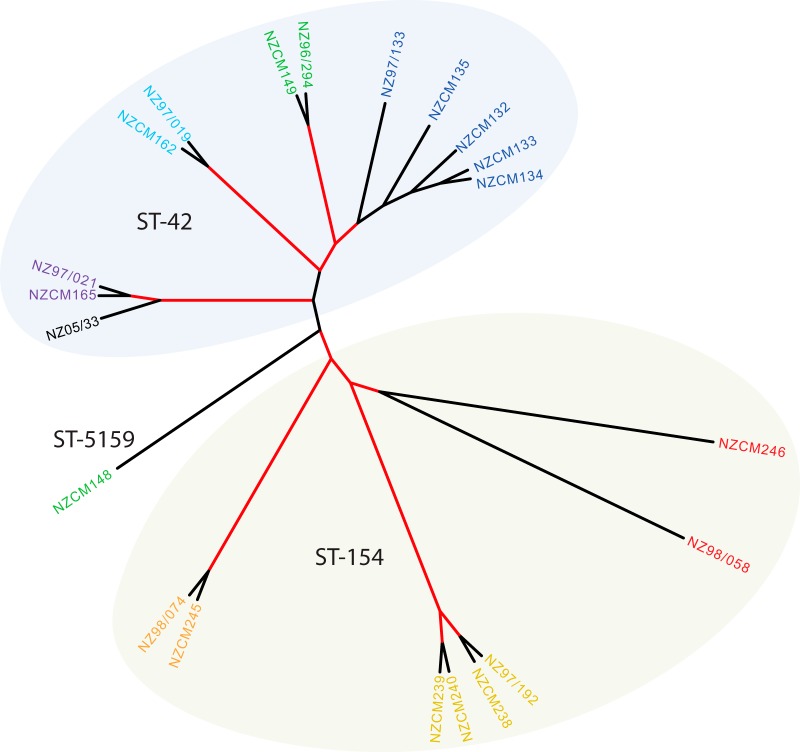
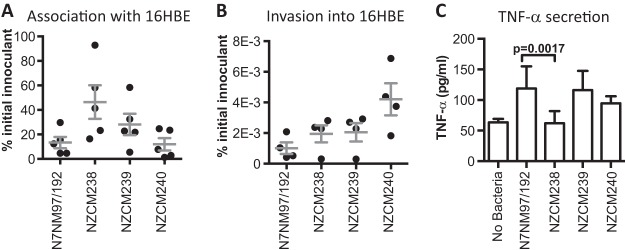
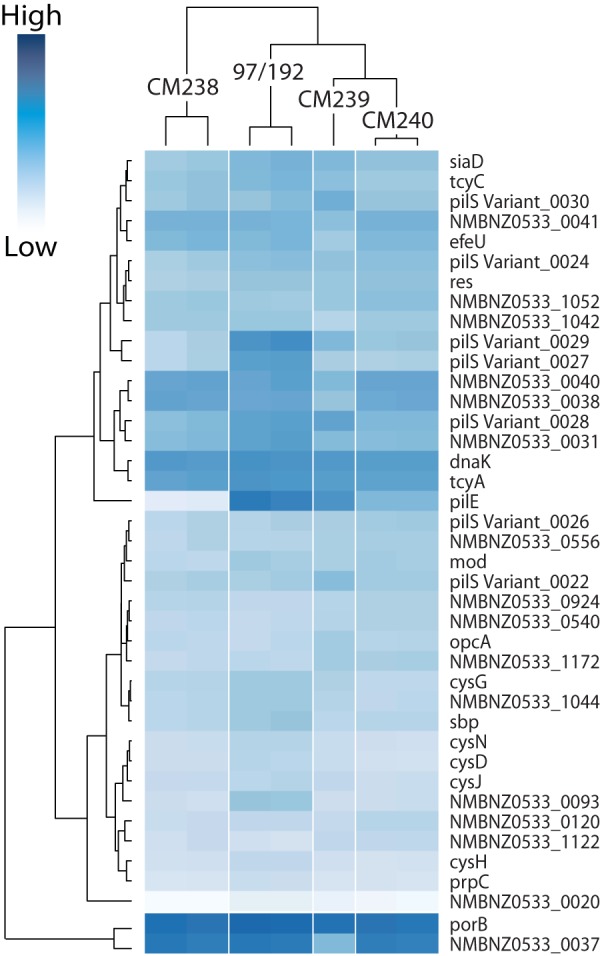
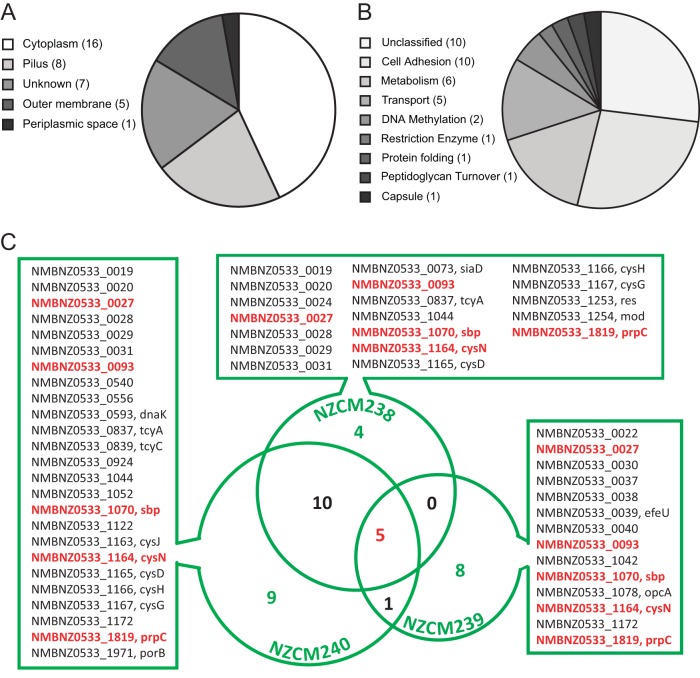
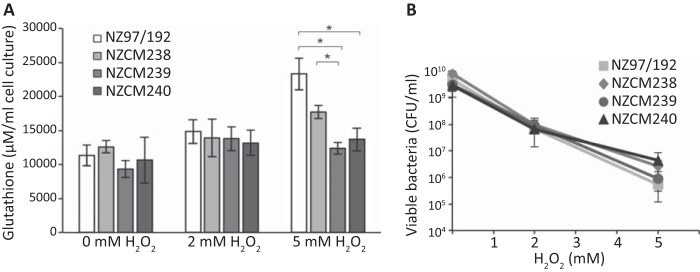
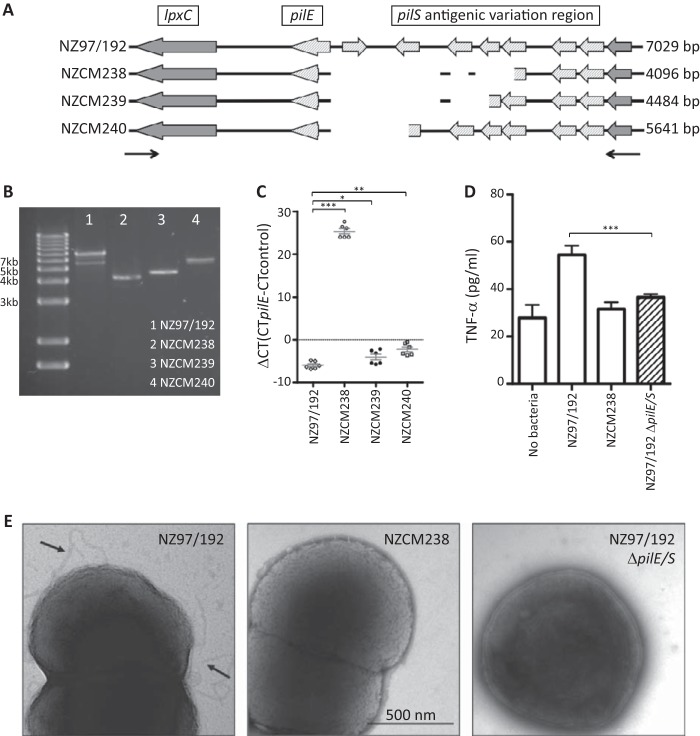
Neisseria meningitidis (meningococcus) can cause meningococcal disease, a rapidly progressing and often fatal disease that can occur in previously healthy children. Meningococci are found in healthy carriers, where they reside in the nasopharynx as commensals. While carriage is relatively common, invasive disease, associated with hypervirulent strains, is a comparatively rare event. The basis of increased virulence in some strains is not well understood. New Zealand suffered a protracted meningococcal disease epidemic, from 1991 to 2008. During this time, a household carriage study was carried out in Auckland: household contacts of index meningococcal disease patients were swabbed for isolation of carriage strains. In many households, healthy carriers harbored strains identical, as determined by laboratory typing, to the ones infecting the associated patient. We carried out more-detailed analyses of carriage and disease isolates from a select number of households. We found that isolates, although indistinguishable by laboratory typing methods and likely closely related, had many differences. We identified multiple genome variants and transcriptional differences between isolates. These studies enabled the identification of two new phase-variable genes. We also found that several carriage strains had lost their type IV pili and that this loss correlated with reduced tumor necrosis factor alpha (TNF-α) expression when cultured with epithelial cells. While nonpiliated meningococcal isolates have been previously found in carriage strains, this is the first evidence of an association between type IV pili from meningococci and a proinflammatory epithelial response. We also identified potentially important metabolic differences between carriage and disease isolates, including the sulfate assimilation pathway. IMPORTANCENeisseria meningitidis causes meningococcal disease but is frequently carried in the throats of healthy individuals; the factors that determine whether invasive disease develops are not completely understood. We carried out detailed studies of isolates, collected from patients and their household contacts, to identify differences between commensal throat isolates and those that caused invasive disease. Though isolates were identical by laboratory typing methods, we uncovered many differences in their genomes, in gene expression, and in their interactions with host cells. In particular, we found that several carriage isolates had lost their type IV pili, a surprising finding since pili are often described as essential for colonization. However, loss of type IV pili correlated with reduced secretion of a proinflammatory cytokine, TNF-α, when meningococci were cocultured with human bronchial epithelial cells; hence, the loss of pili could provide an advantage to meningococci, by resulting in a dampened localized host immune response.
Keywords: Neisseria meningitidis; carriage; household contact; type IV pili.
Figures
Similar articles
-
Attachment of Neisseria meningitidis to human mucosal surfaces: influence of pili and type of receptor cell.J Infect Dis. 1981 Apr;143(4):525-32. doi: 10.1093/infdis/143.4.525. J Infect Dis. 1981. PMID: 6113259
-
Meningococcal carriage in close contacts of cases.Epidemiol Infect. 1991 Feb;106(1):133-41. doi: 10.1017/s0950268800056491. Epidemiol Infect. 1991. PMID: 1899646 Free PMC article.
-
Disease-associated Neisseria meningitidis isolates inhibit wound repair in respiratory epithelial cells in a type IV pilus-independent manner.Infect Immun. 2014 Dec;82(12):5023-34. doi: 10.1128/IAI.02001-14. Epub 2014 Sep 15. Infect Immun. 2014. PMID: 25225250 Free PMC article.
-
Neisseria meningitidis: an overview of the carriage state.J Med Microbiol. 2004 Sep;53(Pt 9):821-832. doi: 10.1099/jmm.0.45529-0. J Med Microbiol. 2004. PMID: 15314188 Review.
-
An unwanted guest: Neisseria meningitidis - carriage, risk for invasive disease and the impact of vaccination with insight on Italy incidence.Expert Rev Anti Infect Ther. 2017 Jul;15(7):689-701. doi: 10.1080/14787210.2017.1333422. Epub 2017 May 29. Expert Rev Anti Infect Ther. 2017. PMID: 28524748 Review.
Cited by
-
Acquisition of virulence genes by a carrier strain gave rise to the ongoing epidemics of meningococcal disease in West Africa.Proc Natl Acad Sci U S A. 2018 May 22;115(21):5510-5515. doi: 10.1073/pnas.1802298115. Epub 2018 May 7. Proc Natl Acad Sci U S A. 2018. PMID: 29735685 Free PMC article.
-
Neisseria meningitidis: using genomics to understand diversity, evolution and pathogenesis.Nat Rev Microbiol. 2020 Feb;18(2):84-96. doi: 10.1038/s41579-019-0282-6. Epub 2019 Nov 8. Nat Rev Microbiol. 2020. PMID: 31705134 Review.
References
-
- Caugant DA, Frøholm LO, Bøvre K, Holten E, Frasch CE, Mocca LF, Zollinger WD, Selander RK. 1986. Intercontinental spread of a genetically distinctive complex of clones of Neisseria meningitidis causing epidemic disease. Proc Natl Acad Sci U S A 83:4927–4931. doi:10.1073/pnas.83.13.4927. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
