

Tumor Necrosis Factor-Mediated Survival of CD169+ Cells Promotes Immune Activation during Vesicular Stomatitis Virus Infection
- PMID: 29142134
- PMCID: PMC5774891
- DOI: 10.1128/JVI.01637-17
Tumor Necrosis Factor-Mediated Survival of CD169+ Cells Promotes Immune Activation during Vesicular Stomatitis Virus Infection
Abstract
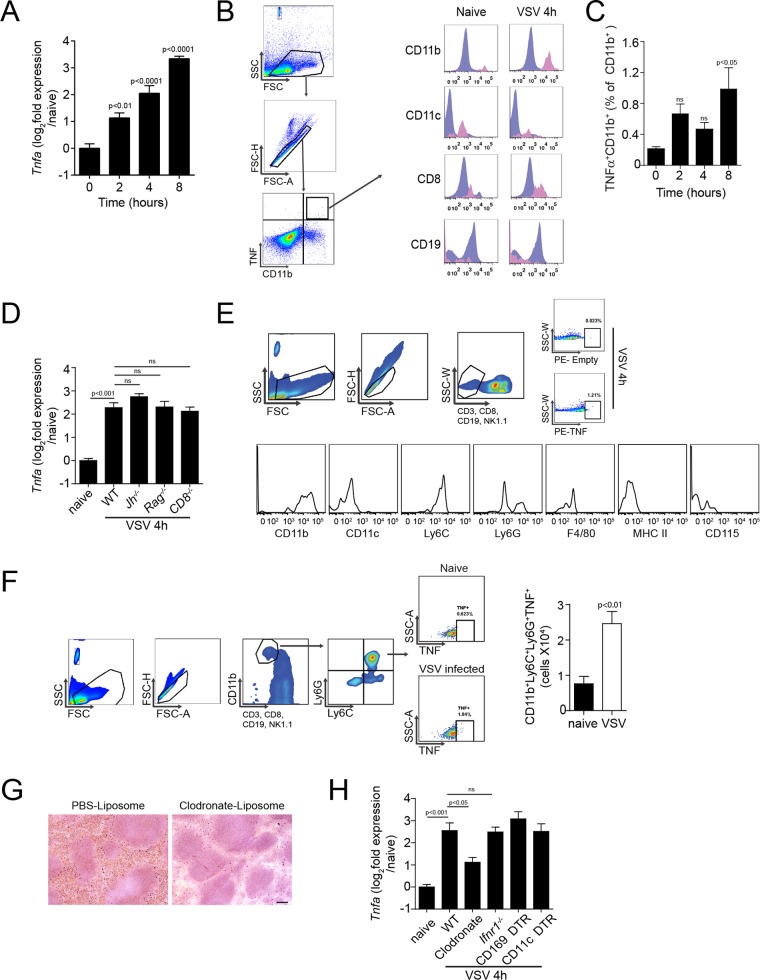
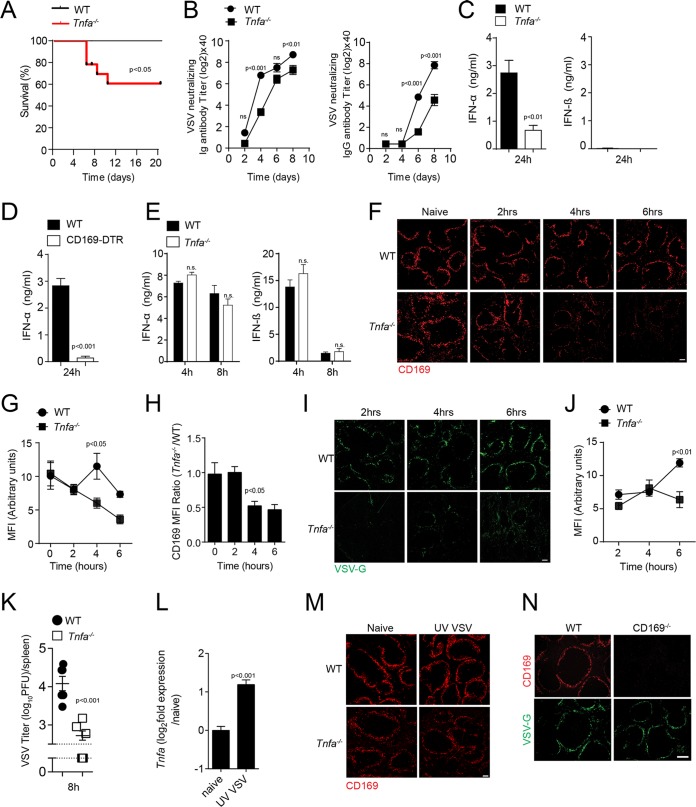
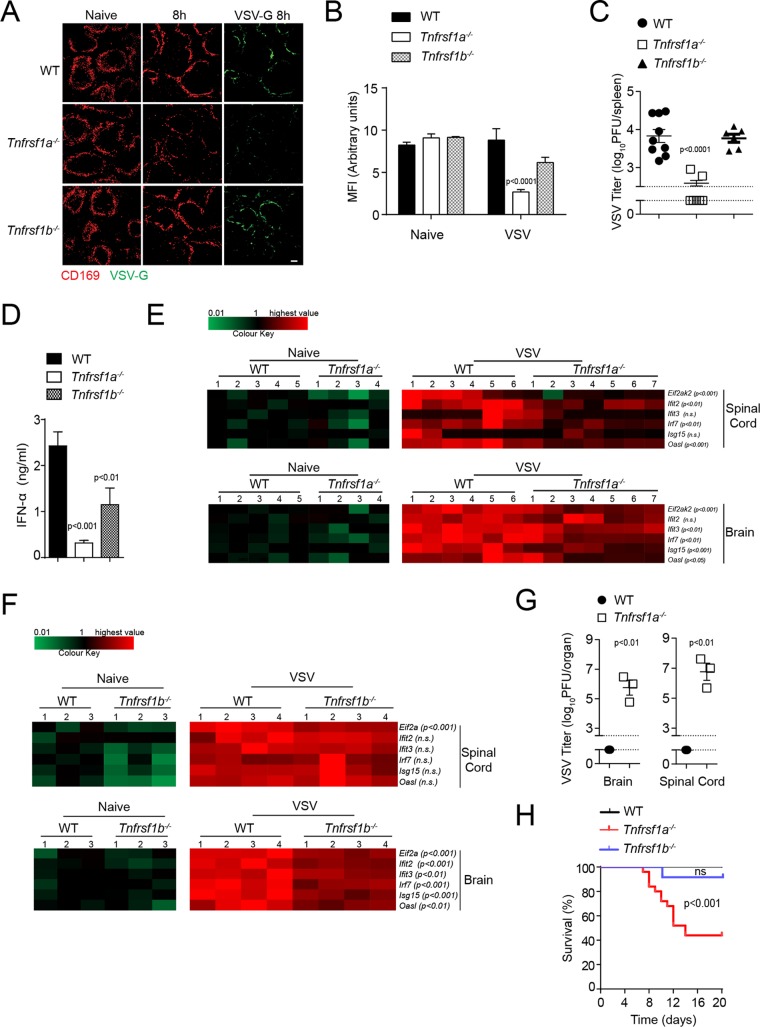
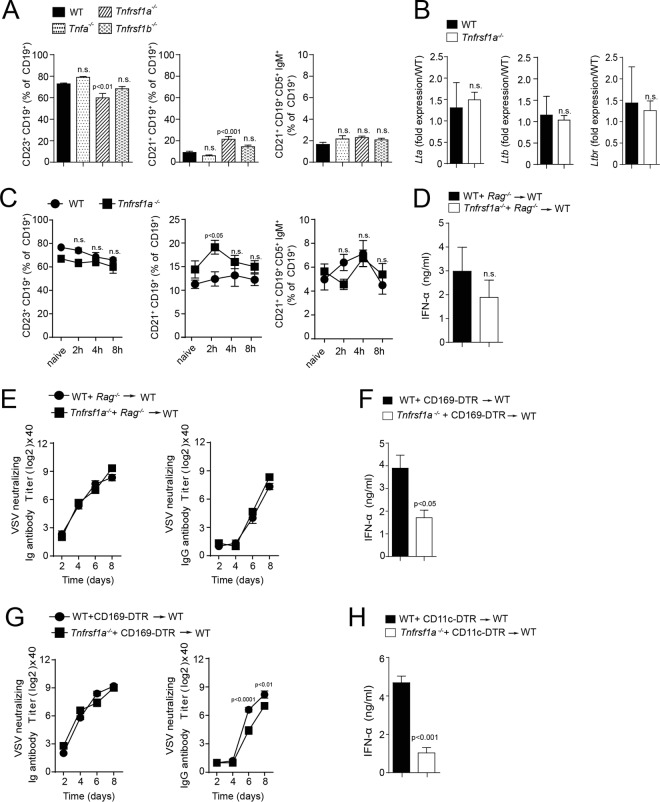
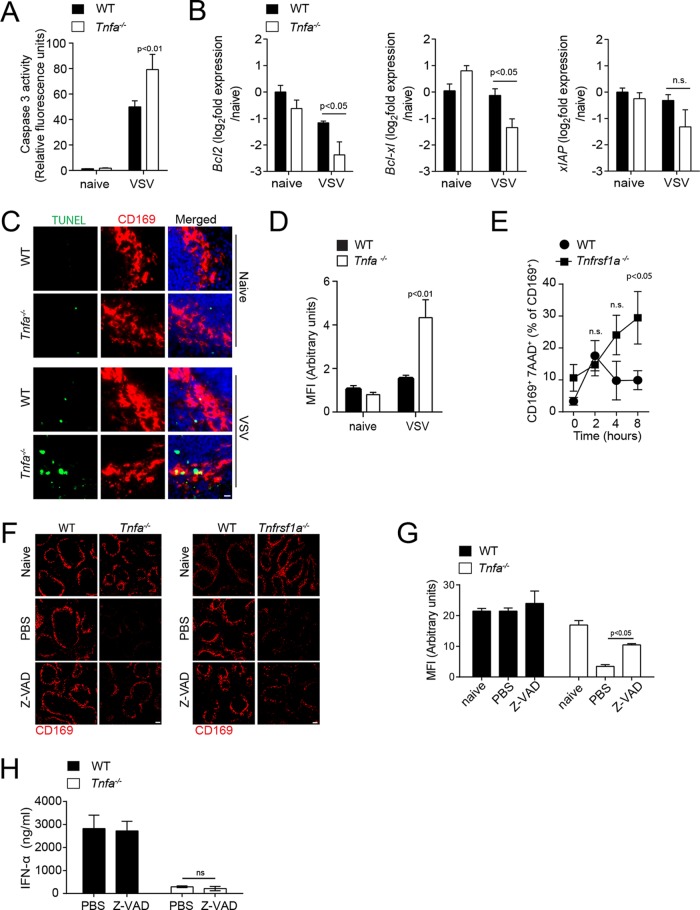
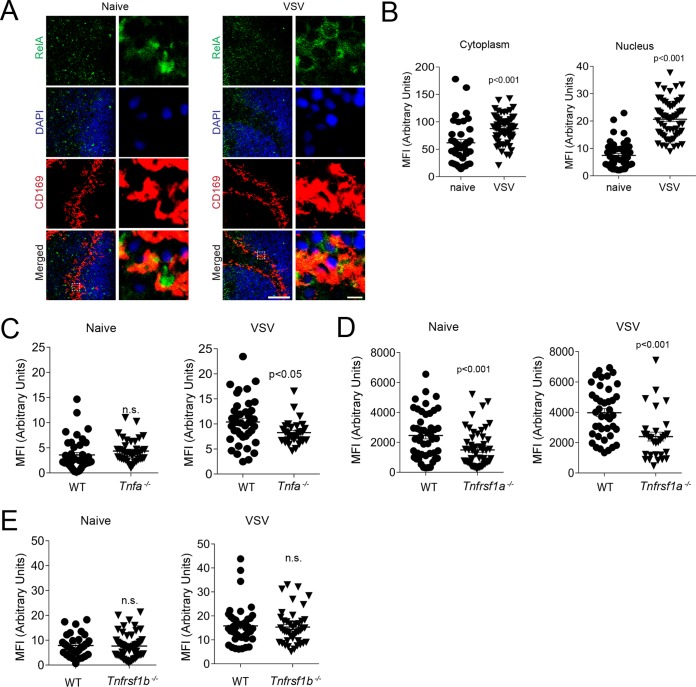
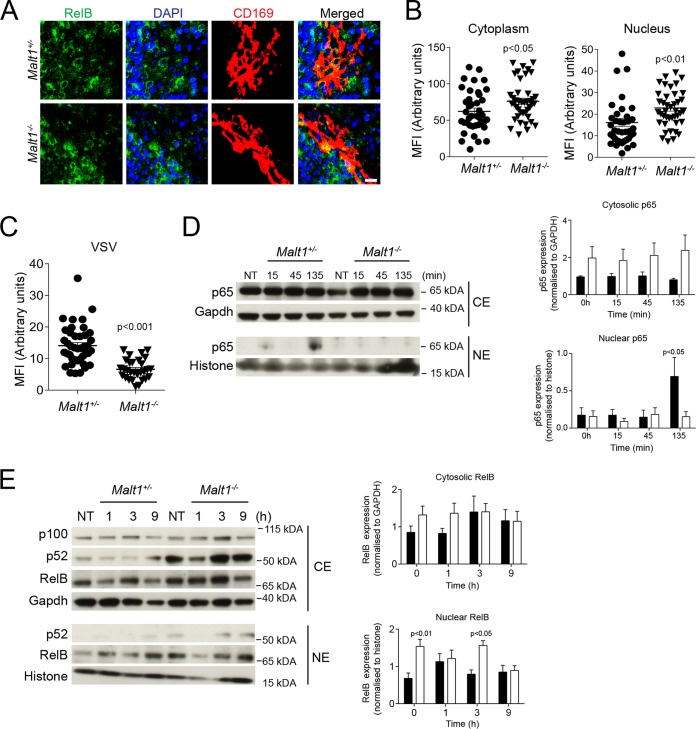
Innate immune activation is essential to mount an effective antiviral response and to prime adaptive immunity. Although a crucial role of CD169+ cells during vesicular stomatitis virus (VSV) infections is increasingly recognized, factors regulating CD169+ cells during viral infections remain unclear. Here, we show that tumor necrosis factor is produced by CD11b+ Ly6C+ Ly6G+ cells following infection with VSV. The absence of TNF or TNF receptor 1 (TNFR1) resulted in reduced numbers of CD169+ cells and in reduced type I interferon (IFN-I) production during VSV infection, with a severe disease outcome. Specifically, TNF triggered RelA translocation into the nuclei of CD169+ cells; this translocation was inhibited when the paracaspase MALT-1 was absent. Consequently, MALT1 deficiency resulted in reduced VSV replication, defective innate immune activation, and development of severe disease. These findings indicate that TNF mediates the maintenance of CD169+ cells and innate and adaptive immune activation during VSV infection.IMPORTANCE Over the last decade, strategically placed CD169+ metallophilic macrophages in the marginal zone of the murine spleen and lymph nodes (LN) have been shown to play a very important role in host defense against viral pathogens. CD169+ macrophages have been shown to activate innate and adaptive immunity via "enforced virus replication," a controlled amplification of virus particles. However, the factors regulating the CD169+ macrophages remain to be studied. In this paper, we show that after vesicular stomatitis virus infection, phagocytes produce tumor necrosis factor (TNF), which signals via TNFR1, and promote enforced virus replication in CD169+ macrophages. Consequently, lack of TNF or TNFR1 resulted in defective immune activation and VSV clearance.
Keywords: MALT1; NF-κB; TNF; innate immunity; interferon; interferons; tumor necrosis factor.
Copyright © 2018 Shinde et al.
Figures

Similar articles
-
Deficiency of the B cell-activating factor receptor results in limited CD169+ macrophage function during viral infection.J Virol. 2015 May;89(9):4748-59. doi: 10.1128/JVI.02976-14. Epub 2015 Feb 11. J Virol. 2015. PMID: 25673724 Free PMC article.
-
Immunoactivation induced by chronic viral infection inhibits viral replication and drives immunosuppression through sustained IFN-I responses.Eur J Immunol. 2016 Feb;46(2):372-80. doi: 10.1002/eji.201545765. Epub 2015 Nov 17. Eur J Immunol. 2016. PMID: 26507703 Free PMC article.
-
Too much of a good thing: Sustained type 1 interferon signaling limits humoral responses to secondary viral infection.Eur J Immunol. 2016 Feb;46(2):300-2. doi: 10.1002/eji.201546224. Eur J Immunol. 2016. PMID: 26783074 Free PMC article.
-
Mechanisms of lymphatic system-specific viral replication and its potential role in autoimmune disease.Clin Exp Immunol. 2019 Jan;195(1):64-73. doi: 10.1111/cei.13241. Clin Exp Immunol. 2019. PMID: 30444956 Free PMC article. Review.
-
CD169+ Macrophages Capture and Dendritic Cells Instruct: The Interplay of the Gatekeeper and the General of the Immune System.Front Immunol. 2018 Oct 26;9:2472. doi: 10.3389/fimmu.2018.02472. eCollection 2018. Front Immunol. 2018. PMID: 30416504 Free PMC article. Review.
Cited by
-
Myeloid CD169/Siglec1: An immunoregulatory biomarker in viral disease.Front Med (Lausanne). 2022 Sep 23;9:979373. doi: 10.3389/fmed.2022.979373. eCollection 2022. Front Med (Lausanne). 2022. PMID: 36213653 Free PMC article. Review.
-
Macrophage Depletion Mitigates Platelet Aggregate Formation in Splenic Marginal Zone and Alleviates LPS-Associated Thrombocytopenia in Rats.Front Med (Lausanne). 2019 Dec 17;6:300. doi: 10.3389/fmed.2019.00300. eCollection 2019. Front Med (Lausanne). 2019. PMID: 31921873 Free PMC article.
-
Innate immune surveillance of the circulation: A review on the removal of circulating virions from the bloodstream.PLoS Pathog. 2022 May 5;18(5):e1010474. doi: 10.1371/journal.ppat.1010474. eCollection 2022 May. PLoS Pathog. 2022. PMID: 35511797 Free PMC article. Review.
-
Functions of CD169 positive macrophages in human diseases (Review).Biomed Rep. 2021 Feb;14(2):26. doi: 10.3892/br.2020.1402. Epub 2020 Dec 17. Biomed Rep. 2021. PMID: 33408860 Free PMC article. Review.
-
Validation of a Quick Flow Cytometry-Based Assay for Acute Infection Based on CD64 and CD169 Expression. New Tools for Early Diagnosis in COVID-19 Pandemic.Front Med (Lausanne). 2021 Mar 23;8:655785. doi: 10.3389/fmed.2021.655785. eCollection 2021. Front Med (Lausanne). 2021. PMID: 33869256 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
