

Global analysis of ribosome-associated noncoding RNAs unveils new modes of translational regulation
- PMID: 29087317
- PMCID: PMC5699049
- DOI: 10.1073/pnas.1708433114
Global analysis of ribosome-associated noncoding RNAs unveils new modes of translational regulation
Abstract
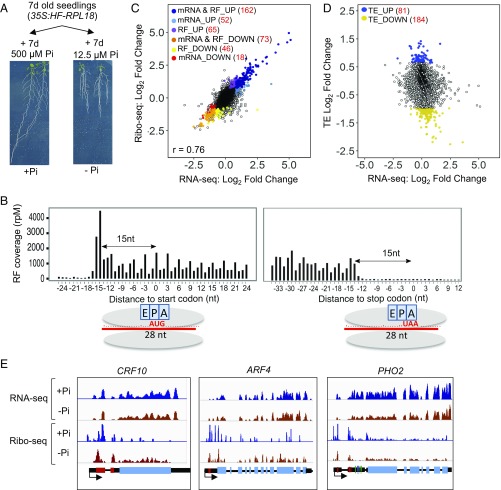
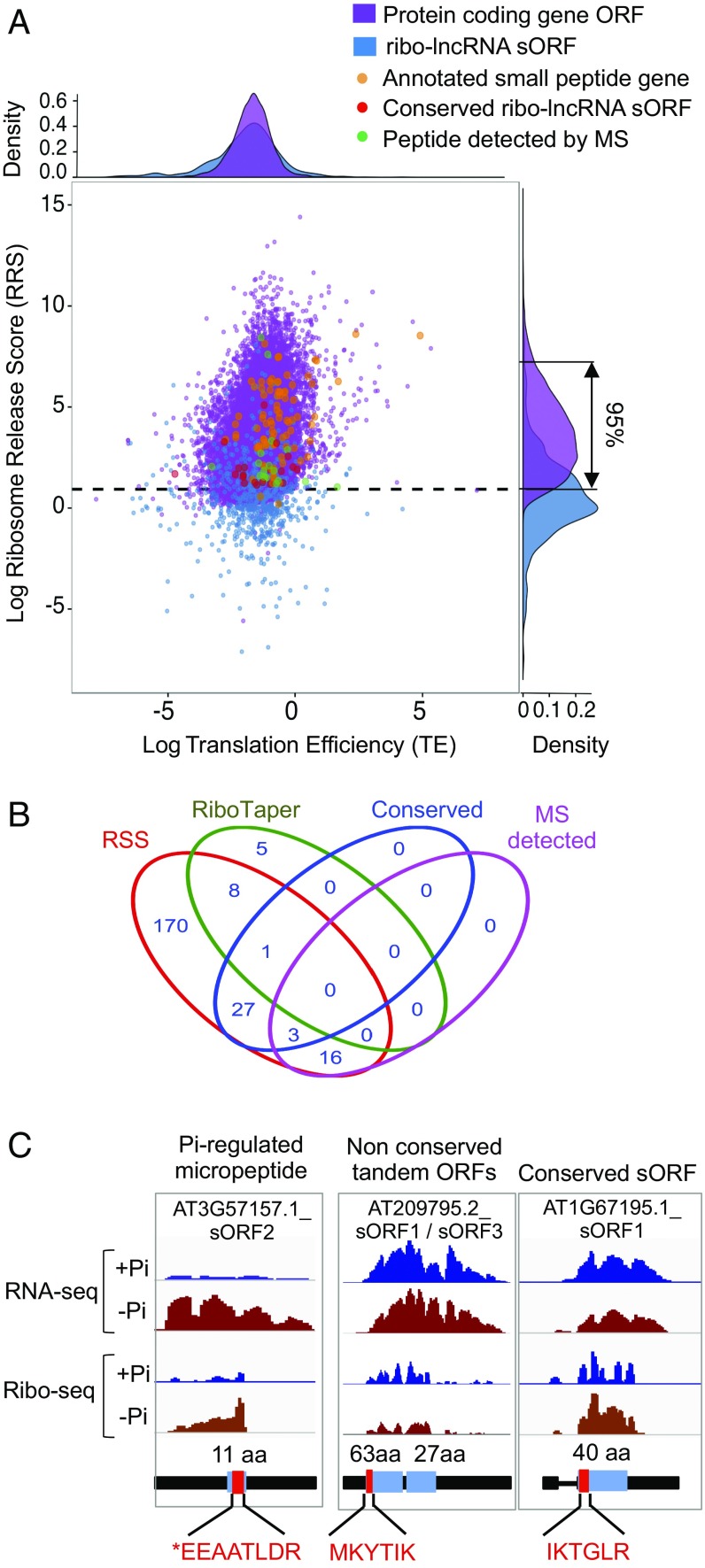
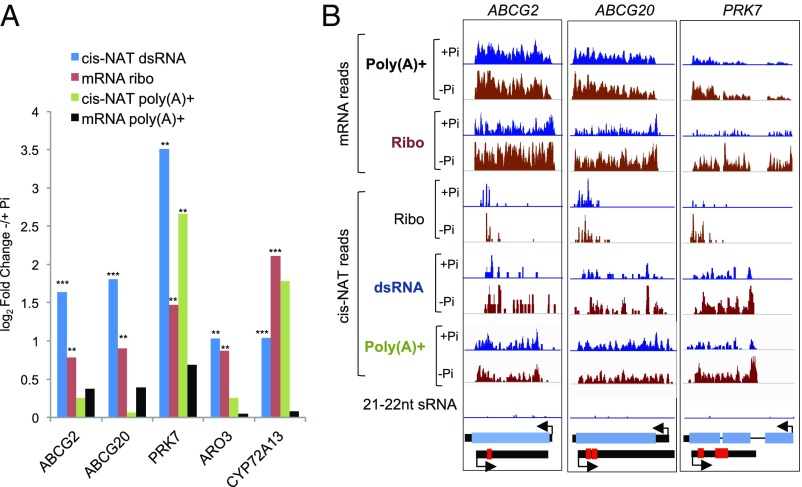
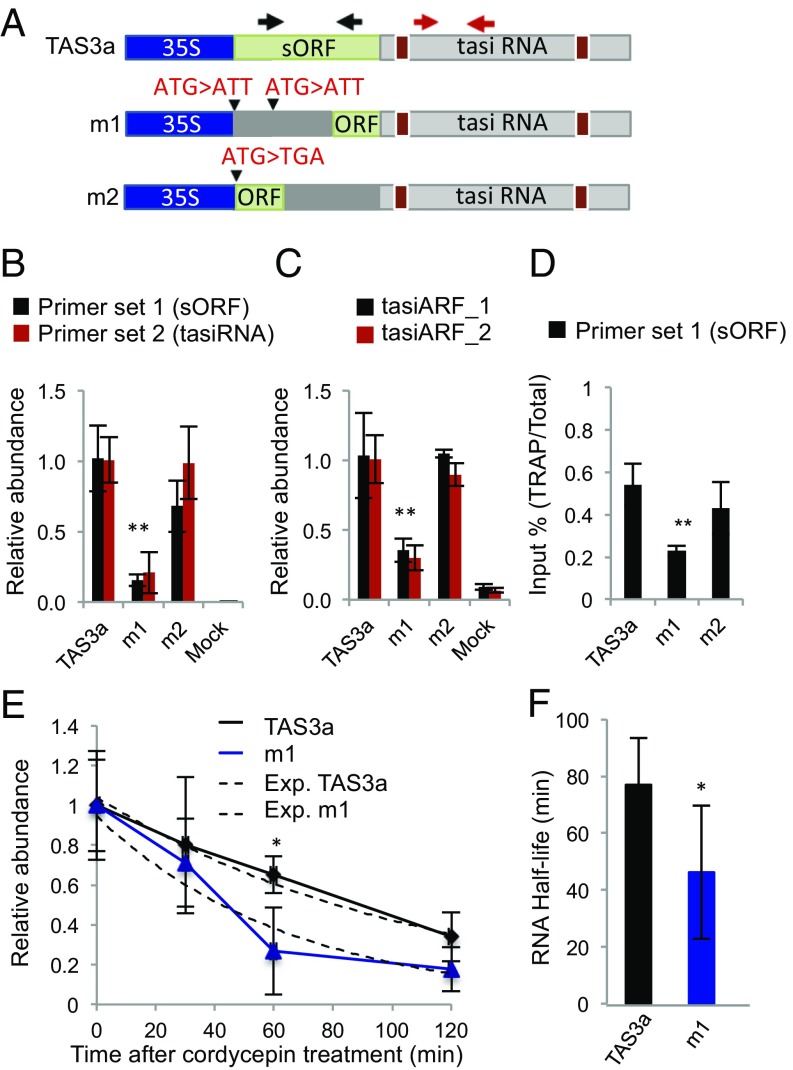
Eukaryotic transcriptomes contain a major non-protein-coding component that includes precursors of small RNAs as well as long noncoding RNA (lncRNAs). Here, we utilized the mapping of ribosome footprints on RNAs to explore translational regulation of coding and noncoding RNAs in roots of Arabidopsis thaliana shifted from replete to deficient phosphorous (Pi) nutrition. Homodirectional changes in steady-state mRNA abundance and translation were observed for all but 265 annotated protein-coding genes. Of the translationally regulated mRNAs, 30% had one or more upstream ORF (uORF) that influenced the number of ribosomes on the principal protein-coding region. Nearly one-half of the 2,382 lncRNAs detected had ribosome footprints, including 56 with significantly altered translation under Pi-limited nutrition. The prediction of translated small ORFs (sORFs) by quantitation of translation termination and peptidic analysis identified lncRNAs that produce peptides, including several deeply evolutionarily conserved and significantly Pi-regulated lncRNAs. Furthermore, we discovered that natural antisense transcripts (NATs) frequently have actively translated sORFs, including five with low-Pi up-regulation that correlated with enhanced translation of the sense protein-coding mRNA. The data also confirmed translation of miRNA target mimics and lncRNAs that produce trans-acting or phased small-interfering RNA (tasiRNA/phasiRNAs). Mutational analyses of the positionally conserved sORF of TAS3a linked its translation with tasiRNA biogenesis. Altogether, this systematic analysis of ribosome-associated mRNAs and lncRNAs demonstrates that nutrient availability and translational regulation controls protein and small peptide-encoding mRNAs as well as a diverse cadre of regulatory RNAs.
Keywords: Arabidopsis thaliana; long noncoding RNA; phosphate deficiency; ribosome footprint profiling; small peptides.
Copyright © 2017 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Translational dynamics revealed by genome-wide profiling of ribosome footprints in Arabidopsis.Proc Natl Acad Sci U S A. 2014 Jan 7;111(1):E203-12. doi: 10.1073/pnas.1317811111. Epub 2013 Dec 23. Proc Natl Acad Sci U S A. 2014. PMID: 24367078 Free PMC article.
-
Translational Landscape of Protein-Coding and Non-Protein-Coding RNAs upon Light Exposure in Arabidopsis.Plant Cell Physiol. 2020 Mar 1;61(3):536-545. doi: 10.1093/pcp/pcz219. Plant Cell Physiol. 2020. PMID: 31794029
-
Pateamine A-sensitive ribosome profiling reveals the scope of translation in mouse embryonic stem cells.BMC Genomics. 2016 Jan 14;17:52. doi: 10.1186/s12864-016-2384-0. BMC Genomics. 2016. PMID: 26764022 Free PMC article.
-
Evolution of new proteins from translated sORFs in long non-coding RNAs.Exp Cell Res. 2020 Jun 1;391(1):111940. doi: 10.1016/j.yexcr.2020.111940. Epub 2020 Mar 7. Exp Cell Res. 2020. PMID: 32156600 Review.
-
Ribosomal profiling adds new coding sequences to the proteome.Biochem Soc Trans. 2015 Dec;43(6):1271-6. doi: 10.1042/BST20150170. Biochem Soc Trans. 2015. PMID: 26614672 Review.
Cited by
-
Beyond transcription: compelling open questions in plant RNA biology.Plant Cell. 2023 May 29;35(6):1626-1653. doi: 10.1093/plcell/koac346. Plant Cell. 2023. PMID: 36477566 Free PMC article.
-
Ribosome-Associated ncRNAs (rancRNAs) Adjust Translation and Shape Proteomes.Noncoding RNA. 2022 Mar 10;8(2):22. doi: 10.3390/ncrna8020022. Noncoding RNA. 2022. PMID: 35314615 Free PMC article. Review.
-
Plant Long Noncoding RNAs: New Players in the Field of Post-Transcriptional Regulations.Noncoding RNA. 2021 Feb 17;7(1):12. doi: 10.3390/ncrna7010012. Noncoding RNA. 2021. PMID: 33671131 Free PMC article. Review.
-
An Evolutionarily Conserved AU-Rich Element in the 3' Untranslated Region of a Transcript Misannotated as a Long Noncoding RNA Regulates RNA Stability.Mol Cell Biol. 2022 Apr 21;42(4):e0050521. doi: 10.1128/mcb.00505-21. Epub 2022 Mar 11. Mol Cell Biol. 2022. PMID: 35274990 Free PMC article.
-
Plants utilise ancient conserved peptide upstream open reading frames in stress-responsive translational regulation.Plant Cell Environ. 2022 Apr;45(4):1229-1241. doi: 10.1111/pce.14277. Epub 2022 Feb 15. Plant Cell Environ. 2022. PMID: 35128674 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
