

Vitamin D as a Novel Regulator of Tumor Metabolism: Insights on Potential Mechanisms and Implications for Anti-Cancer Therapy
- PMID: 29048387
- PMCID: PMC5666865
- DOI: 10.3390/ijms18102184
Vitamin D as a Novel Regulator of Tumor Metabolism: Insights on Potential Mechanisms and Implications for Anti-Cancer Therapy
Abstract
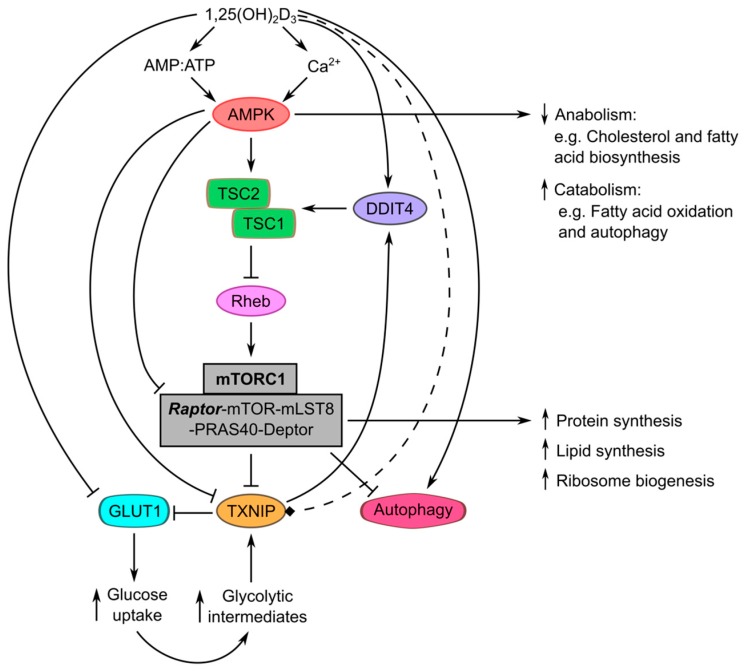
1,25-dihydroxyvitamin D₃ [1,25(OH)₂D₃], the bioactive form of vitamin D, has been shown to possess significant anti-tumor potential. While most studies so far have focused on the ability of this molecule to influence the proliferation and apoptosis of cancer cells, more recent data indicate that 1,25(OH)₂D₃ also impacts energy utilization in tumor cells. In this article, we summarize and review the evidence that demonstrates the targeting of metabolic aberrations in cancers by 1,25(OH)₂D₃, and highlight potential mechanisms through which these effects may be executed. We shed light on the ability of this molecule to regulate metabolism-related tumor suppressors and oncogenes, energy- and nutrient-sensing pathways, as well as cell death and survival mechanisms such as autophagy.
Keywords: AMPK (AMP-activated protein kinase); HIF1a (Hypoxia-inducible factor 1a); TXNIP (Thioredoxin-interacting protein); autophagy; c-Myc; cancer; mTOR (Mammalian target of rapamycin); metabolism; p53; vitamin D.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
A novel cytostatic form of autophagy in sensitization of non-small cell lung cancer cells to radiation by vitamin D and the vitamin D analog, EB 1089.Autophagy. 2014;10(12):2346-61. doi: 10.4161/15548627.2014.993283. Autophagy. 2014. PMID: 25629933 Free PMC article.
-
Differences in p53 status significantly influence the cellular response and cell survival to 1,25-dihydroxyvitamin D3-metformin cotreatment in colorectal cancer cells.Mol Carcinog. 2017 Nov;56(11):2486-2498. doi: 10.1002/mc.22696. Epub 2017 Jul 13. Mol Carcinog. 2017. PMID: 28618116
-
Antiproliferative effects of 1alpha,25-dihydroxyvitamin D(3) and vitamin D analogs on tumor-derived endothelial cells.Endocrinology. 2002 Jul;143(7):2508-14. doi: 10.1210/endo.143.7.8887. Endocrinology. 2002. PMID: 12072382
-
Molecular aspects of vitamin D anticancer activity.Cancer Invest. 2012 Oct;30(8):604-14. doi: 10.3109/07357907.2012.721039. Epub 2012 Sep 10. Cancer Invest. 2012. PMID: 22963190 Review.
-
Hypoxia-mediated drug resistance: novel insights on the functional interaction of HIFs and cell death pathways.Drug Resist Updat. 2011 Jun;14(3):191-201. doi: 10.1016/j.drup.2011.03.001. Epub 2011 Apr 3. Drug Resist Updat. 2011. PMID: 21466972 Review.
Cited by
-
Immunomodulatory Role of Thioredoxin Interacting Protein in Cancer's Impediments: Current Understanding and Therapeutic Implications.Vaccines (Basel). 2022 Nov 10;10(11):1902. doi: 10.3390/vaccines10111902. Vaccines (Basel). 2022. PMID: 36366411 Free PMC article. Review.
-
Antitumor effects and mechanisms of 1,25(OH)2D3 in the Pfeiffer diffuse large B lymphoma cell line.Mol Med Rep. 2019 Dec;20(6):5064-5074. doi: 10.3892/mmr.2019.10756. Epub 2019 Oct 17. Mol Med Rep. 2019. PMID: 31638226 Free PMC article.
-
Evaluation of the p53 pathway in polycystic ovarian syndrome pathogenesis and apoptosis enhancement in human granulosa cells through transcriptome data analysis.Sci Rep. 2023 Jul 19;13(1):11648. doi: 10.1038/s41598-023-38340-1. Sci Rep. 2023. PMID: 37468508 Free PMC article.
-
Understanding the Biological Activities of Vitamin D in Type 1 Neurofibromatosis: New Insights into Disease Pathogenesis and Therapeutic Design.Cancers (Basel). 2020 Oct 13;12(10):2965. doi: 10.3390/cancers12102965. Cancers (Basel). 2020. PMID: 33066259 Free PMC article. Review.
-
Vitamin D and Hypoxia: Points of Interplay in Cancer.Cancers (Basel). 2022 Mar 31;14(7):1791. doi: 10.3390/cancers14071791. Cancers (Basel). 2022. PMID: 35406562 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
