

Zdhhc13-dependent Drp1 S-palmitoylation impacts brain bioenergetics, anxiety, coordination and motor skills
- PMID: 29038583
- PMCID: PMC5643561
- DOI: 10.1038/s41598-017-12889-0
Zdhhc13-dependent Drp1 S-palmitoylation impacts brain bioenergetics, anxiety, coordination and motor skills
Abstract
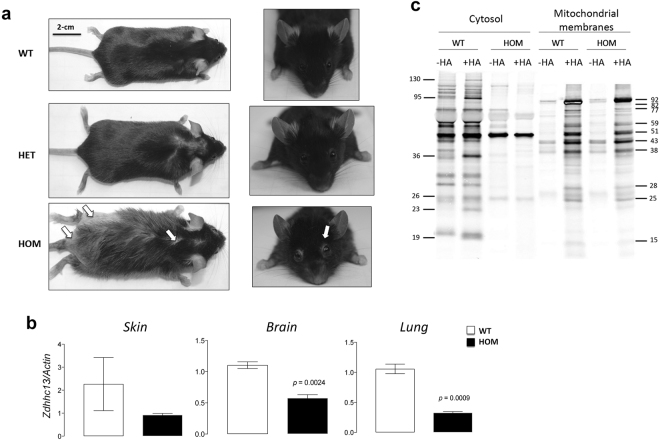
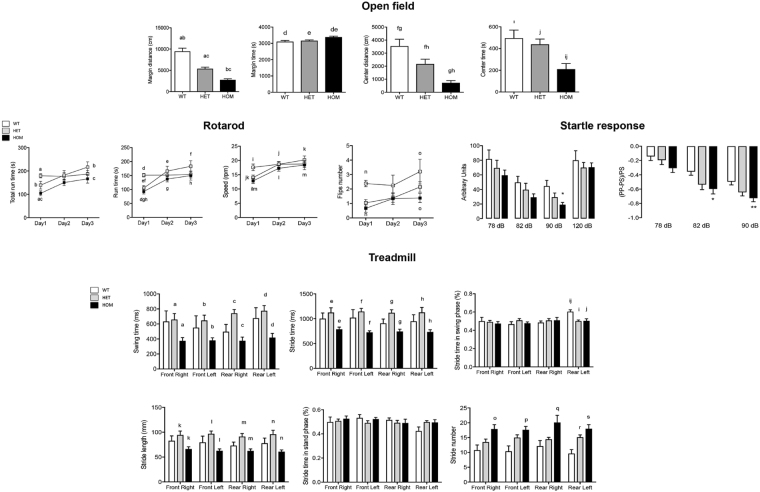
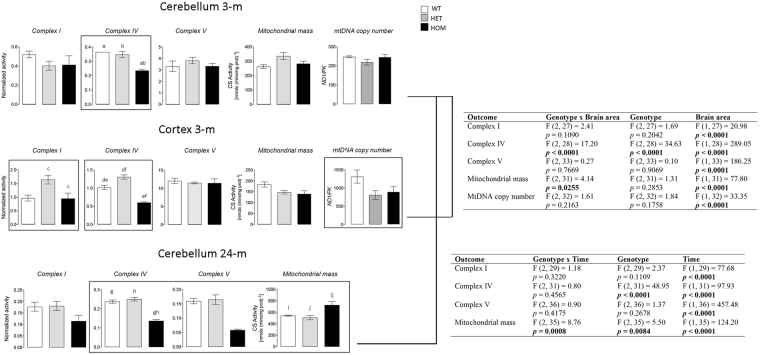
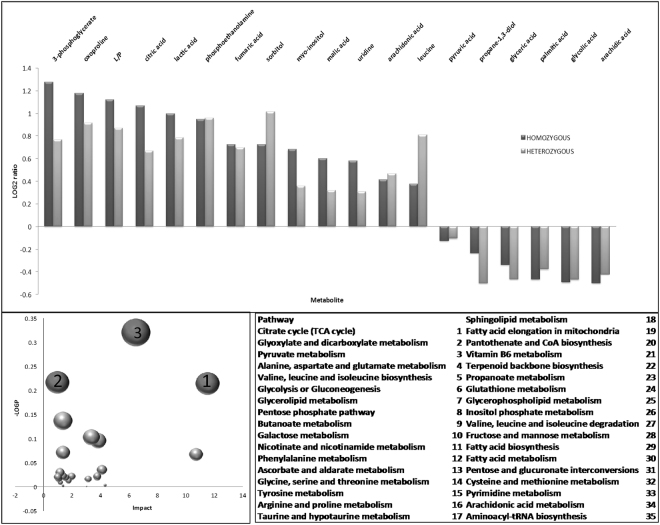
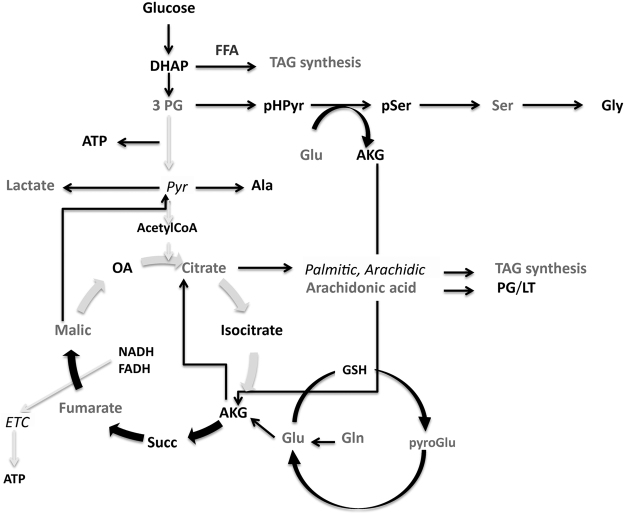
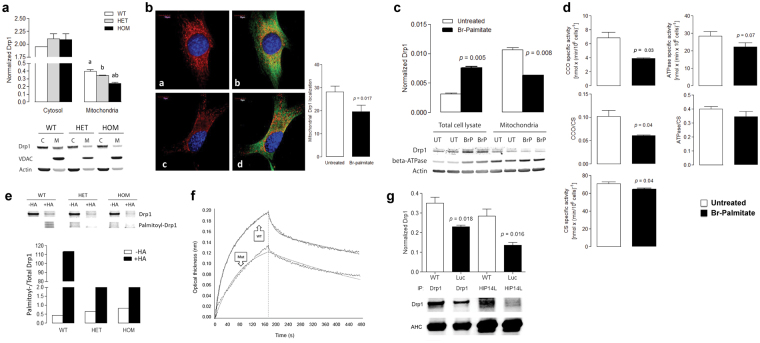
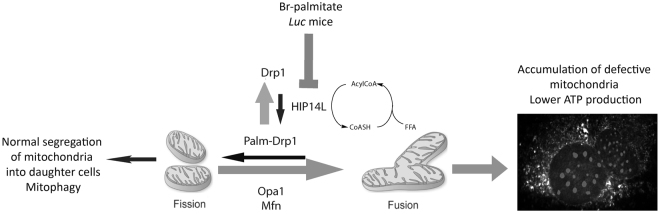
Protein S-palmitoylation is a reversible post-translational modification mediated by palmitoyl acyltransferase enzymes, a group of Zn2+-finger DHHC-domain-containing proteins (ZDHHC). Here, for the first time, we show that Zdhhc13 plays a key role in anxiety-related behaviors and motor function, as well as brain bioenergetics, in a mouse model (luc) carrying a spontaneous Zdhhc13 recessive mutation. At 3 m of age, mutant mice displayed increased sensorimotor gating, anxiety, hypoactivity, and decreased motor coordination, compared to littermate controls. Loss of Zdhhc13 in cortex and cerebellum from 3- and 24 m old hetero- and homozygous male mutant mice resulted in lower levels of Drp1 S-palmitoylation accompanied by altered mitochondrial dynamics, increased glycolysis, glutaminolysis and lactic acidosis, and neurotransmitter imbalances. Employing in vivo and in vitro models, we identified that Zdhhc13-dependent Drp1 S-palmitoylation, which acting alone or in concert, enables the normal occurrence of the fission-fusion process. In vitro and in vivo direct Zdhhc13-Drp1 protein interaction was observed, confirming Drp1 as a substrate of Zdhhc13. Abnormal fission-fusion processes result in disrupted mitochondria morphology and distribution affecting not only mitochondrial ATP output but neurotransmission and integrity of synaptic structures in the brain, setting the basis for the behavioral abnormalities described in the Zdhhc13-deficient mice.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Palmitoyl acyltransferase, Zdhhc13, facilitates bone mass acquisition by regulating postnatal epiphyseal development and endochondral ossification: a mouse model.PLoS One. 2014 Mar 17;9(3):e92194. doi: 10.1371/journal.pone.0092194. eCollection 2014. PLoS One. 2014. PMID: 24637783 Free PMC article.
-
Palmitoyl Acyltransferase Activity of ZDHHC13 Regulates Skin Barrier Development Partly by Controlling PADi3 and TGM1 Protein Stability.J Invest Dermatol. 2020 May;140(5):959-970.e3. doi: 10.1016/j.jid.2019.09.017. Epub 2019 Oct 25. J Invest Dermatol. 2020. PMID: 31669413
-
Circadian Control of DRP1 Activity Regulates Mitochondrial Dynamics and Bioenergetics.Cell Metab. 2018 Mar 6;27(3):657-666.e5. doi: 10.1016/j.cmet.2018.01.011. Epub 2018 Feb 22. Cell Metab. 2018. PMID: 29478834
-
The ZDHHC13/ZDHHC17 subfamily: From biological functions to therapeutic targets of diseases.Pharmacol Res. 2024 Nov;209:107418. doi: 10.1016/j.phrs.2024.107418. Epub 2024 Sep 19. Pharmacol Res. 2024. PMID: 39306022 Review.
-
Emerging Roles of DHHC-mediated Protein S-palmitoylation in Physiological and Pathophysiological Context.Eur J Cell Biol. 2018 Jun;97(5):319-338. doi: 10.1016/j.ejcb.2018.03.005. Epub 2018 Mar 22. Eur J Cell Biol. 2018. PMID: 29602512 Review.
Cited by
-
Protein posttranslational modifications in health and diseases: Functions, regulatory mechanisms, and therapeutic implications.MedComm (2020). 2023 May 2;4(3):e261. doi: 10.1002/mco2.261. eCollection 2023 Jun. MedComm (2020). 2023. PMID: 37143582 Free PMC article. Review.
-
AMPK Phosphorylates ZDHHC13 to Increase MC1R Activity and Suppress Melanomagenesis.Cancer Res. 2023 Apr 4;83(7):1062-1073. doi: 10.1158/0008-5472.CAN-22-2595. Cancer Res. 2023. PMID: 36701140 Free PMC article.
-
Post-translational palmitoylation of metabolic proteins.Front Physiol. 2023 Feb 24;14:1122895. doi: 10.3389/fphys.2023.1122895. eCollection 2023. Front Physiol. 2023. PMID: 36909239 Free PMC article. Review.
-
2-Bromopalmitate attenuates bone cancer pain via reversing mitochondrial fusion and fission imbalance in spinal astrocytes.Mol Pain. 2019 Jan-Dec;15:1744806919871813. doi: 10.1177/1744806919871813. Mol Pain. 2019. PMID: 31394961 Free PMC article.
-
Hepatic palmitoyl-proteomes and acyl-protein thioesterase protein proximity networks link lipid modification and mitochondria.Cell Rep. 2023 Nov 28;42(11):113389. doi: 10.1016/j.celrep.2023.113389. Epub 2023 Nov 4. Cell Rep. 2023. PMID: 37925639 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
