

Natural Reassortants of Potentially Zoonotic Avian Influenza Viruses H5N1 and H9N2 from Egypt Display Distinct Pathogenic Phenotypes in Experimentally Infected Chickens and Ferrets
- PMID: 28931674
- PMCID: PMC5686759
- DOI: 10.1128/JVI.01300-17
Natural Reassortants of Potentially Zoonotic Avian Influenza Viruses H5N1 and H9N2 from Egypt Display Distinct Pathogenic Phenotypes in Experimentally Infected Chickens and Ferrets
Abstract
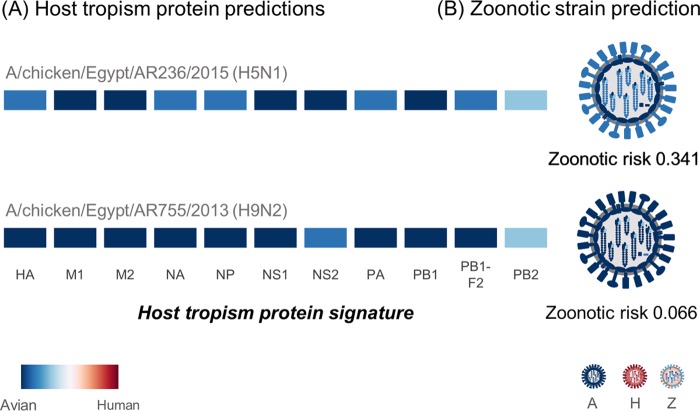
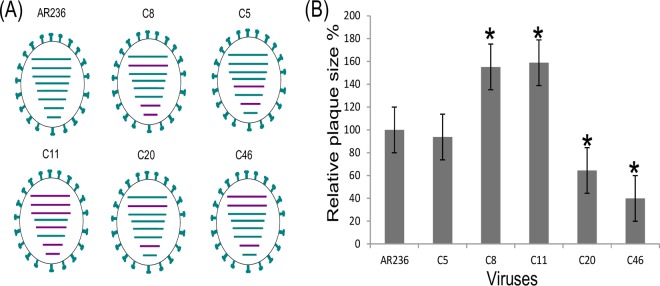
The cocirculation of zoonotic highly pathogenic avian influenza virus (HPAIV) of subtype H5N1 and avian influenza virus (AIV) of subtype H9N2 among poultry in Egypt for at least 6 years should render that country a hypothetical hot spot for the emergence of reassortant, phenotypically altered viruses, yet no reassortants have been detected in Egypt. The present investigations proved that reassortants of the Egyptian H5N1 clade 2.2.1.2 virus and H9N2 virus of the G1-B lineage can be generated by coamplification in embryonated chicken eggs. Reassortants were restricted to the H5N1 subtype and acquired between two and all six of the internal segments of the H9N2 virus. Five selected plaque-purified reassortant clones expressed a broad phenotypic spectrum both in vitro and in vivo Two groups of reassortants were characterized to have retarded growth characteristics in vitro compared to the H5N1 parent virus. One clone provoked reduced mortality in inoculated chickens, although the characteristics of a highly pathogenic phenotype were retained. Enhanced zoonotic properties were not predicted for any of these clones, and this prediction was confirmed by ferret inoculation experiments: neither the H5N1 parent virus nor two selected clones induced severe clinical symptoms or were transmitted to sentinel ferrets by contact. While the emergence of reassortants of Egyptian HPAIV of subtype H5N1 with internal gene segments of cocirculating H9N2 viruses is possible in principle, the spread of such viruses is expected to be governed by their fitness to outcompete the parental viruses in the field. The eventual spread of attenuated phenotypes, however, would negatively impact syndrome surveillance on poultry farms and might foster enzootic virus circulation.IMPORTANCE Despite almost 6 years of the continuous cocirculation of highly pathogenic avian influenza virus H5N1 and avian influenza virus H9N2 in poultry in Egypt, no reassortants of the two subtypes have been reported. Here, the principal compatibility of the two subtypes is shown by forcing the reassortment between copassaged H5N1 und H9N2 viruses in embryonated chicken eggs. The resulting reassortant viruses displayed a wide range of pathogenicity including attenuated phenotypes in chickens, but did not show enhanced zoonotic propensities in the ferret model.
Keywords: Egypt; highly pathogenic avian influenza; reassortment; viral fitness; zoonosis.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Genetic Compatibility of Reassortants between Avian H5N1 and H9N2 Influenza Viruses with Higher Pathogenicity in Mammals.J Virol. 2019 Feb 5;93(4):e01969-18. doi: 10.1128/JVI.01969-18. Print 2019 Feb 15. J Virol. 2019. PMID: 30463961 Free PMC article.
-
Single gene reassortment of highly pathogenic avian influenza A H5N1 in the low pathogenic H9N2 backbone and its impact on pathogenicity and infectivity of novel reassortant viruses.Arch Virol. 2017 Oct;162(10):2959-2969. doi: 10.1007/s00705-017-3434-x. Epub 2017 Jun 15. Arch Virol. 2017. PMID: 28620809
-
PA from a Recent H9N2 (G1-Like) Avian Influenza a Virus (AIV) Strain Carrying Lysine 367 Confers Altered Replication Efficiency and Pathogenicity to Contemporaneous H5N1 in Mammalian Systems.Viruses. 2020 Sep 20;12(9):1046. doi: 10.3390/v12091046. Viruses. 2020. PMID: 32962203 Free PMC article.
-
Challenge for One Health: Co-Circulation of Zoonotic H5N1 and H9N2 Avian Influenza Viruses in Egypt.Viruses. 2018 Mar 9;10(3):121. doi: 10.3390/v10030121. Viruses. 2018. PMID: 29522492 Free PMC article. Review.
-
Epidemiology, ecology and gene pool of influenza A virus in Egypt: will Egypt be the epicentre of the next influenza pandemic?Virulence. 2015;6(1):6-18. doi: 10.4161/21505594.2014.992662. Virulence. 2015. PMID: 25635701 Free PMC article. Review.
Cited by
-
A Semiquantitative Scoring System for Histopathological and Immunohistochemical Assessment of Lesions and Tissue Tropism in Avian Influenza.Viruses. 2021 May 9;13(5):868. doi: 10.3390/v13050868. Viruses. 2021. PMID: 34065126 Free PMC article.
-
Molecular pathogenic and host range determinants of reassortant Egyptian low pathogenic avian influenza H9N2 viruses from backyard chicken.Int J Vet Sci Med. 2019 Jul 11;7(1):10-19. doi: 10.1080/23144599.2019.1637046. eCollection 2019. Int J Vet Sci Med. 2019. PMID: 31620483 Free PMC article.
-
Identification of Amino Acid Residues Responsible for Inhibition of Host Gene Expression by Influenza A H9N2 NS1 Targeting of CPSF30.Front Microbiol. 2018 Oct 24;9:2546. doi: 10.3389/fmicb.2018.02546. eCollection 2018. Front Microbiol. 2018. PMID: 30405591 Free PMC article.
-
Mass vaccination with reassortment-impaired live H9N2 avian influenza vaccine.NPJ Vaccines. 2024 Aug 3;9(1):136. doi: 10.1038/s41541-024-00923-y. NPJ Vaccines. 2024. PMID: 39097573 Free PMC article.
-
Novel Reassortant Highly Pathogenic Avian Influenza A(H5N2) Virus in Broiler Chickens, Egypt.Emerg Infect Dis. 2020 Jan;26(1):129-133. doi: 10.3201/eid2601.190570. Emerg Infect Dis. 2020. PMID: 31855539 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
