

Understanding the molecular basis for multiple mitochondrial dysfunctions syndrome 1 (MMDS1): impact of a disease-causing Gly189Arg substitution on NFU1
- PMID: 28906594
- PMCID: PMC5696030
- DOI: 10.1111/febs.14271
Understanding the molecular basis for multiple mitochondrial dysfunctions syndrome 1 (MMDS1): impact of a disease-causing Gly189Arg substitution on NFU1
Abstract
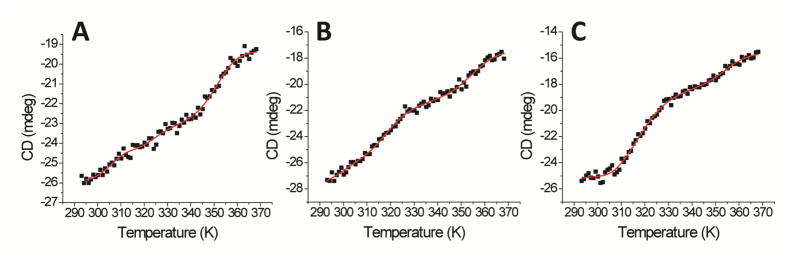
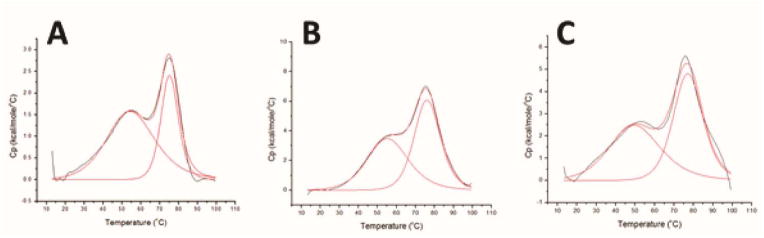
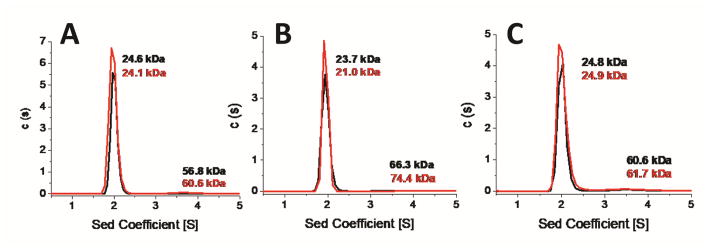
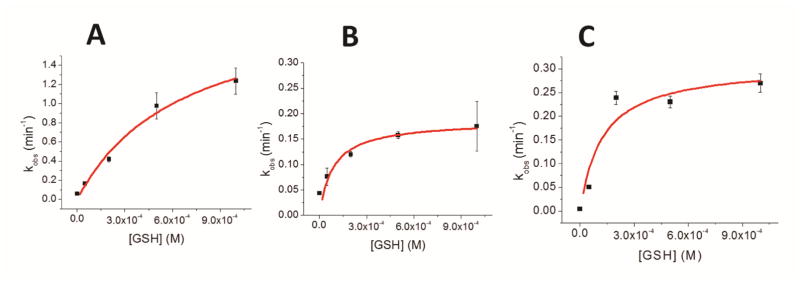
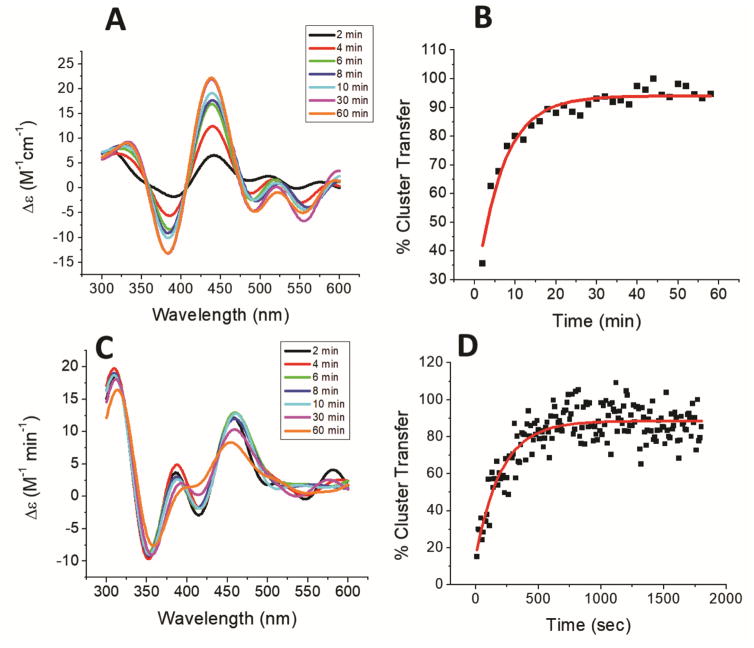
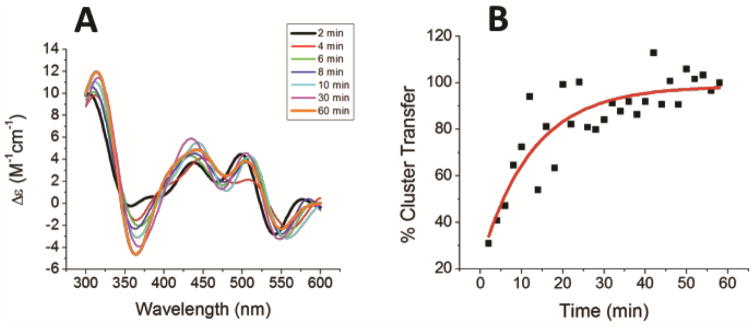
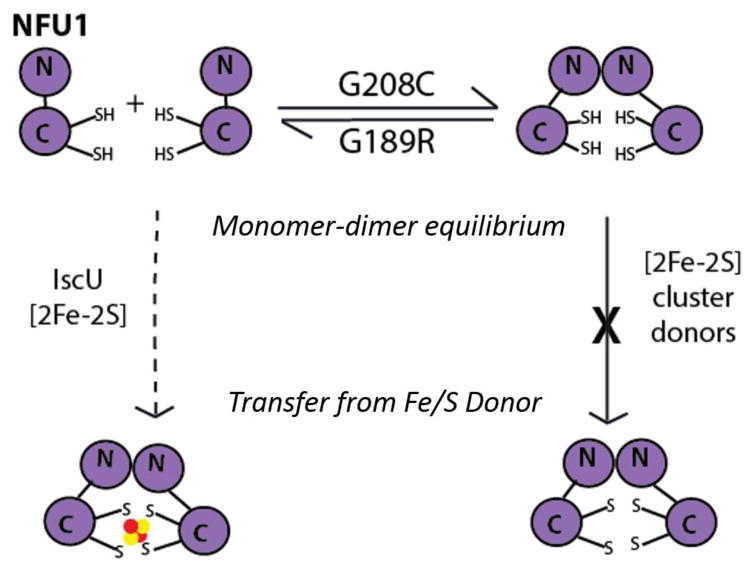
Iron-sulfur (Fe/S) cluster-containing proteins constitute one of the largest protein classes, with highly varied function. Consequently, the biosynthesis of Fe/S clusters is evolutionarily conserved and mutations in intermediate Fe/S cluster scaffold proteins can cause disease, including multiple mitochondrial dysfunctions syndrome (MMDS). Herein, we have characterized the impact of defects occurring in the MMDS1 disease state that result from a point mutation (p.Gly189Arg) near the active site of NFU1, an Fe/S scaffold protein. In vitro investigation into the structure-function relationship of the Gly189Arg derivative, along with two other variants, reveals that substitution at position 189 triggers structural changes that increase flexibility, decrease stability, and alter the monomer-dimer equilibrium toward monomer, thereby impairing the ability of the Gly189X derivatives to receive an Fe/S cluster from physiologically relevant sources.
Keywords: NFU1; cluster exchange; iron-sulfur cluster; mitochondrial disease; protein stability.
© 2017 Federation of European Biochemical Societies.
Figures

Similar articles
-
Analysis of NFU-1 metallocofactor binding-site substitutions-impacts on iron-sulfur cluster coordination and protein structure and function.FEBS J. 2017 Nov;284(22):3817-3837. doi: 10.1111/febs.14270. Epub 2017 Oct 16. FEBS J. 2017. PMID: 28906593
-
Understanding the Molecular Basis of Multiple Mitochondrial Dysfunctions Syndrome 1 (MMDS1)-Impact of a Disease-Causing Gly208Cys Substitution on Structure and Activity of NFU1 in the Fe/S Cluster Biosynthetic Pathway.J Mol Biol. 2017 Mar 24;429(6):790-807. doi: 10.1016/j.jmb.2017.01.021. Epub 2017 Feb 1. J Mol Biol. 2017. PMID: 28161430 Free PMC article.
-
Assembly of the [4Fe-4S] cluster of NFU1 requires the coordinated donation of two [2Fe-2S] clusters from the scaffold proteins, ISCU2 and ISCA1.Hum Mol Genet. 2020 Nov 25;29(19):3165-3182. doi: 10.1093/hmg/ddaa172. Hum Mol Genet. 2020. PMID: 32776106 Free PMC article.
-
Mitochondrial iron-sulfur protein biogenesis and human disease.Biochimie. 2014 May;100:61-77. doi: 10.1016/j.biochi.2014.01.010. Epub 2014 Jan 23. Biochimie. 2014. PMID: 24462711 Review.
-
Posttranslational control of the scaffold for Fe-S cluster biogenesis as a compensatory regulatory mechanism.Curr Genet. 2017 Feb;63(1):51-56. doi: 10.1007/s00294-016-0618-y. Epub 2016 May 31. Curr Genet. 2017. PMID: 27246605 Free PMC article. Review.
Cited by
-
Molecular Basis of Rare Diseases Associated to the Maturation of Mitochondrial [4Fe-4S]-Containing Proteins.Biomolecules. 2022 Jul 21;12(7):1009. doi: 10.3390/biom12071009. Biomolecules. 2022. PMID: 35883565 Free PMC article. Review.
-
Role of the HSPA9/HSC20 chaperone pair in promoting directional human iron-sulfur cluster exchange involving monothiol glutaredoxin 5.J Inorg Biochem. 2018 Jul;184:100-107. doi: 10.1016/j.jinorgbio.2018.04.007. Epub 2018 Apr 11. J Inorg Biochem. 2018. PMID: 29689452 Free PMC article.
-
Systematic analysis of expression profiles and prognostic significance for MMDS-related iron-sulfur proteins in renal clear cell carcinoma.Sci Rep. 2022 Nov 16;12(1):19637. doi: 10.1038/s41598-022-22479-4. Sci Rep. 2022. PMID: 36385109 Free PMC article.
-
In Vitro Demonstration of Human Lipoyl Synthase Catalytic Activity in the Presence of NFU1.ACS Bio Med Chem Au. 2022 Oct 19;2(5):456-468. doi: 10.1021/acsbiomedchemau.2c00020. Epub 2022 Jun 13. ACS Bio Med Chem Au. 2022. PMID: 36281303 Free PMC article.
-
Patient-specific variants of NFU1/NFU-1 disrupt cholinergic signaling in a model of multiple mitochondrial dysfunctions syndrome 1.Dis Model Mech. 2023 Feb 1;16(2):dmm049594. doi: 10.1242/dmm.049594. Epub 2023 Feb 1. Dis Model Mech. 2023. PMID: 36645076 Free PMC article.
References
-
- Wachnowsky C, Wesley NA, Fidai I, Cowan JA. Understanding the molecular basis for Multiple Mitochondrial Dysfunctions Syndrome 1 (MMDS1) - Impact of a disease-causing Gly208Cys substitution on structure and activity of NFU1 in the Fe/S cluster biosynthetic pathway. J Mol Biol. 2017;429:790–807. - PMC - PubMed
-
- Lamm O. Die Differentialgleichung der Ultrazentrifugierung. Ark Mat Astr Fys. 1929:1–4.
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
