

Synthetic Toll-Like Receptor 4 (TLR4) and TLR7 Ligands Work Additively via MyD88 To Induce Protective Antiviral Immunity in Mice
- PMID: 28724768
- PMCID: PMC5599744
- DOI: 10.1128/JVI.01050-17
Synthetic Toll-Like Receptor 4 (TLR4) and TLR7 Ligands Work Additively via MyD88 To Induce Protective Antiviral Immunity in Mice
Abstract
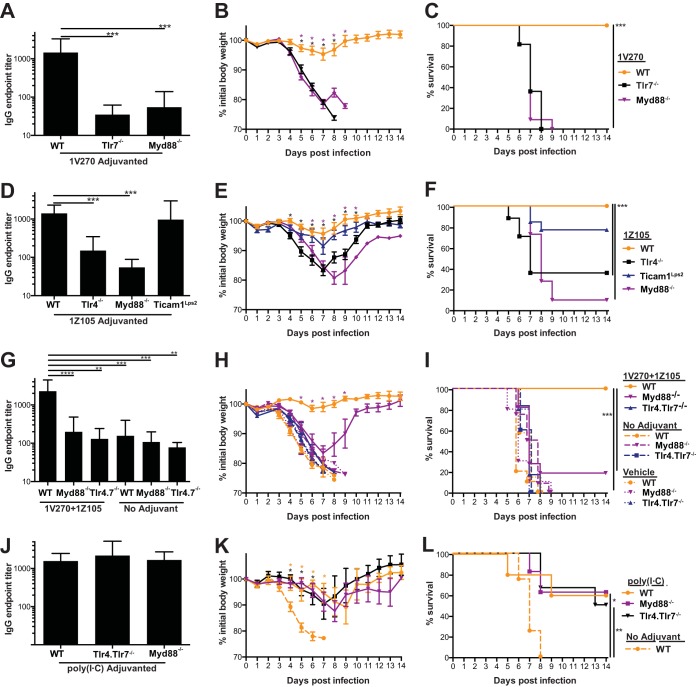
We previously demonstrated that the combination of synthetic small-molecule Toll-like receptor 4 (TLR4) and TLR7 ligands is a potent adjuvant for recombinant influenza virus hemagglutinin, inducing rapid and sustained immunity that is protective against influenza viruses in homologous, heterologous, and heterosubtypic murine challenge models. Combining the TLR4 and TLR7 ligands balances Th1 and Th2-type immune responses for long-lived cellular and neutralizing humoral immunity against the viral hemagglutinin. Here, we demonstrate that the protective response induced in mice by this combined adjuvant is dependent upon TLR4 and TLR7 signaling via myeloid differentiation primary response gene 88 (MyD88), indicating that the adjuvants function in vivo via their known receptors, with negligible off-target effects, to induce protective immunity. The combined adjuvant acts via MyD88 in both bone marrow-derived and non-bone marrow-derived radioresistant cells to induce hemagglutinin-specific antibodies and protect mice against influenza virus challenge. The protective efficacy generated by immunization with this adjuvant and recombinant hemagglutinin antigen is transferable with serum from immunized mice to recipient mice in a homologous, but not a heterologous, H1N1 viral challenge model. Depletion of CD4+ cells after an established humoral response in immunized mice does not impair protection from a homologous challenge; however, it does significantly impair recovery from a heterologous challenge virus, highlighting an important role for vaccine-induced CD4+ cells in cross-protective vaccine efficacy. The combination of the two TLR agonists allows for significant dose reductions of each component to achieve a level of protection equivalent to that afforded by either single agent at its full dose.IMPORTANCE Development of novel adjuvants is needed to enhance immunogenicity to provide better protection from seasonal influenza virus infection and improve pandemic preparedness. We show here that several dose combinations of synthetic TLR4 and TLR7 ligands are potent adjuvants for recombinant influenza virus hemagglutinin antigen induction of humoral and cellular immunity against viral challenges. The components of the combined adjuvant work additively to enable both antigen and adjuvant dose sparing while retaining efficacy. Understanding an adjuvant's mechanism of action is a critical component for preclinical safety evaluation, and we demonstrate here that a combined TLR4 and TLR7 adjuvant signals via the appropriate receptors and the MyD88 adaptor protein. This novel adjuvant combination contributes to a more broadly protective vaccine while demonstrating an attractive safety profile.
Keywords: TLR4; TLR7; Toll-like receptor; adjuvant; influenza virus; vaccine.
Copyright © 2017 American Society for Microbiology.
Figures




Similar articles
-
A Novel Synthetic Dual Agonistic Liposomal TLR4/7 Adjuvant Promotes Broad Immune Responses in an Influenza Vaccine With Minimal Reactogenicity.Front Immunol. 2020 Jun 19;11:1207. doi: 10.3389/fimmu.2020.01207. eCollection 2020. Front Immunol. 2020. PMID: 32636840 Free PMC article.
-
Synthetic Toll-like receptor 4 (TLR4) and TLR7 ligands as influenza virus vaccine adjuvants induce rapid, sustained, and broadly protective responses.J Virol. 2015 Mar;89(6):3221-35. doi: 10.1128/JVI.03337-14. Epub 2015 Jan 7. J Virol. 2015. PMID: 25568203 Free PMC article.
-
TLR7 recognition is dispensable for influenza virus A infection but important for the induction of hemagglutinin-specific antibodies in response to the 2009 pandemic split vaccine in mice.J Virol. 2012 Oct;86(20):10988-98. doi: 10.1128/JVI.01064-12. Epub 2012 Jul 25. J Virol. 2012. PMID: 22837197 Free PMC article.
-
Next-Generation Influenza HA Immunogens and Adjuvants in Pursuit of a Broadly Protective Vaccine.Viruses. 2021 Mar 24;13(4):546. doi: 10.3390/v13040546. Viruses. 2021. PMID: 33805245 Free PMC article. Review.
-
Cross-protective immunity against influenza virus infections induced by intranasal vaccination together with a TLR3-mucosal adjuvant.Hum Vaccin. 2011 Jan-Feb;7 Suppl:174-82. doi: 10.4161/hv.7.0.14584. Epub 2011 Jan 1. Hum Vaccin. 2011. PMID: 21321485 Review.
Cited by
-
MEK1/2 Inhibitors Unlock the Constrained Interferon Response in Macrophages Through IRF1 Signaling.Front Immunol. 2019 Aug 27;10:2020. doi: 10.3389/fimmu.2019.02020. eCollection 2019. Front Immunol. 2019. PMID: 31507609 Free PMC article.
-
The cationic liposome CCS/C adjuvant induces immunity to influenza independently of the adaptor protein MyD88.Hum Vaccin Immunother. 2020 Dec 1;16(12):3146-3154. doi: 10.1080/21645515.2020.1750247. Epub 2020 May 13. Hum Vaccin Immunother. 2020. PMID: 32401698 Free PMC article.
-
Allicin Alleviates Reticuloendotheliosis Virus-Induced Immunosuppression via ERK/Mitogen-Activated Protein Kinase Pathway in Specific Pathogen-Free Chickens.Front Immunol. 2017 Dec 22;8:1856. doi: 10.3389/fimmu.2017.01856. eCollection 2017. Front Immunol. 2017. PMID: 29312337 Free PMC article.
-
A Novel Synthetic Dual Agonistic Liposomal TLR4/7 Adjuvant Promotes Broad Immune Responses in an Influenza Vaccine With Minimal Reactogenicity.Front Immunol. 2020 Jun 19;11:1207. doi: 10.3389/fimmu.2020.01207. eCollection 2020. Front Immunol. 2020. PMID: 32636840 Free PMC article.
-
TLR7 promotes chronic airway disease in RSV-infected mice.Front Immunol. 2023 Sep 14;14:1240552. doi: 10.3389/fimmu.2023.1240552. eCollection 2023. Front Immunol. 2023. PMID: 37795093 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
