

Jasmonate-mediated defence responses, unlike salicylate-mediated responses, are involved in the recovery of grapevine from bois noir disease
- PMID: 28693415
- PMCID: PMC5504844
- DOI: 10.1186/s12870-017-1069-4
Jasmonate-mediated defence responses, unlike salicylate-mediated responses, are involved in the recovery of grapevine from bois noir disease
Erratum in
-
Correction to: Jasmonate-mediated defence responses, unlike salicylate-mediated responses, are involved in the recovery of grapevine from bois noir disease.BMC Plant Biol. 2018 Feb 21;18(1):39. doi: 10.1186/s12870-018-1251-3. BMC Plant Biol. 2018. PMID: 29466943 Free PMC article.
Abstract
Background: Bois noir is an important disease of grapevine (Vitis vinifera L.), caused by phytoplasmas. An interesting, yet elusive aspect of the bois noir disease is "recovery", i.e., the spontaneous and unpredictable remission of symptoms and damage. Because conventional pest management is ineffective against bois noir, deciphering the molecular bases of recovery is beneficial. The present study aimed to understand whether salicylate- and jasmonate-defence pathways might have a role in the recovery from the bois noir disease of grapevine.
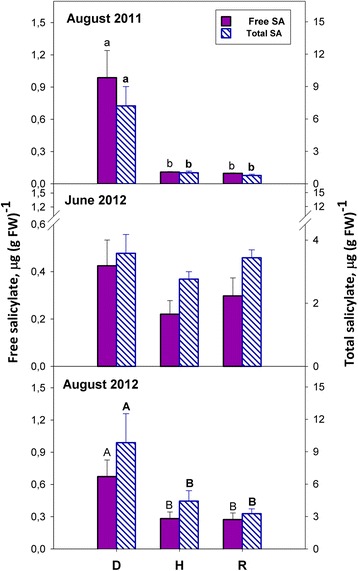
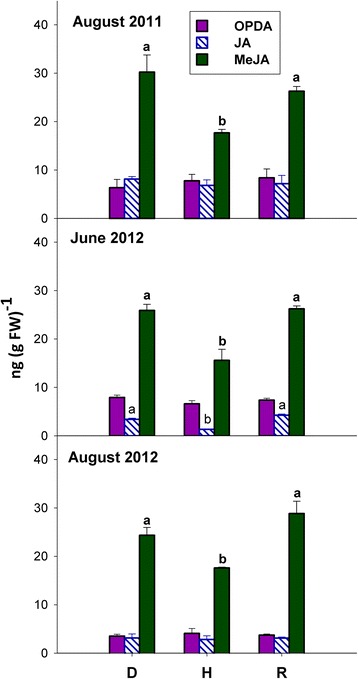
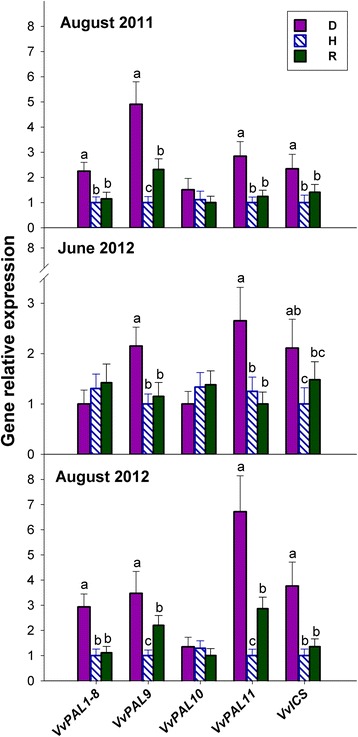
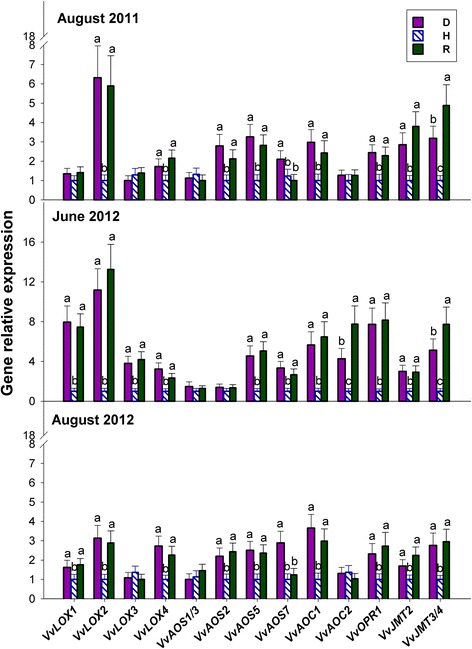
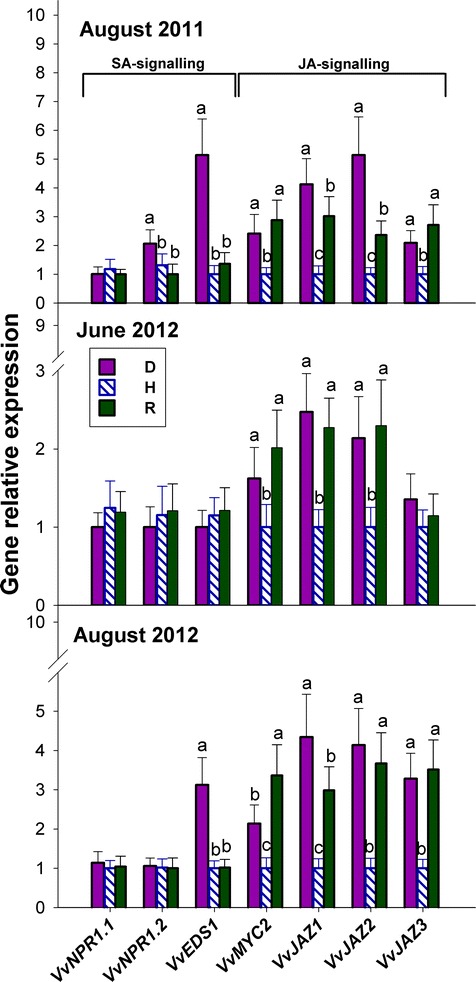
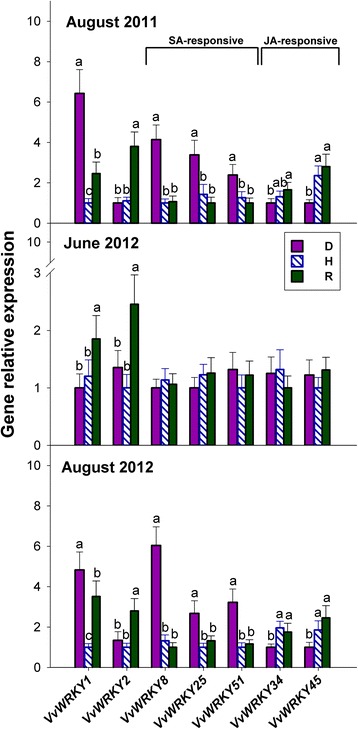
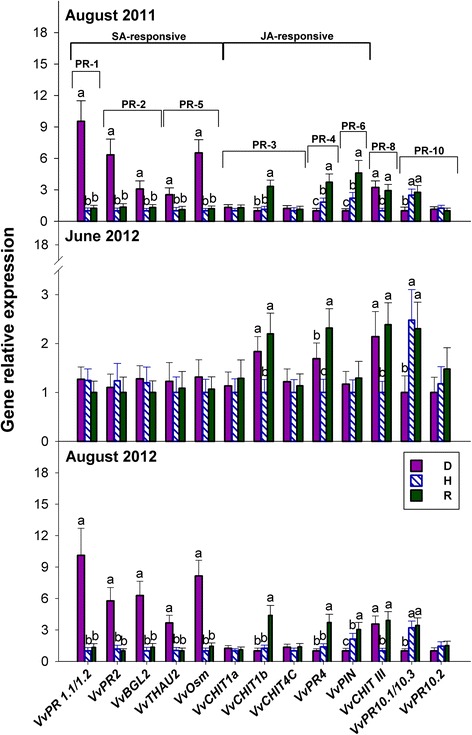
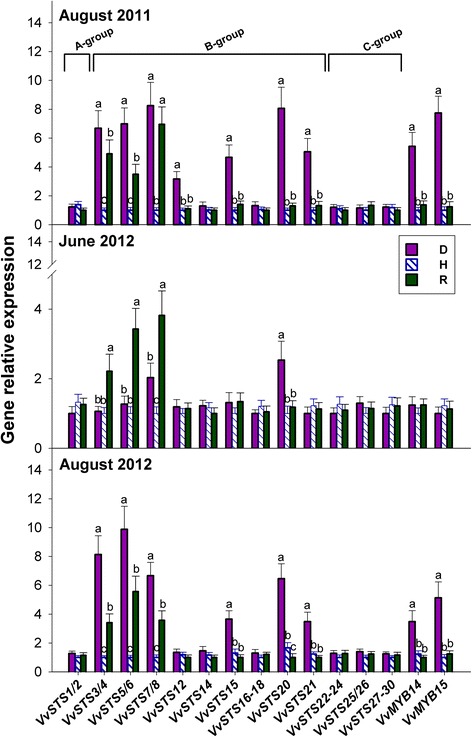
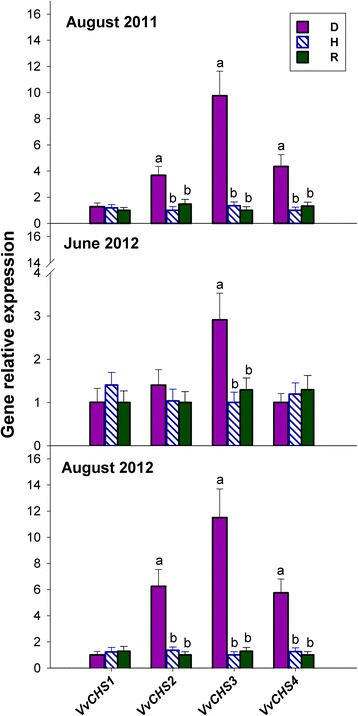
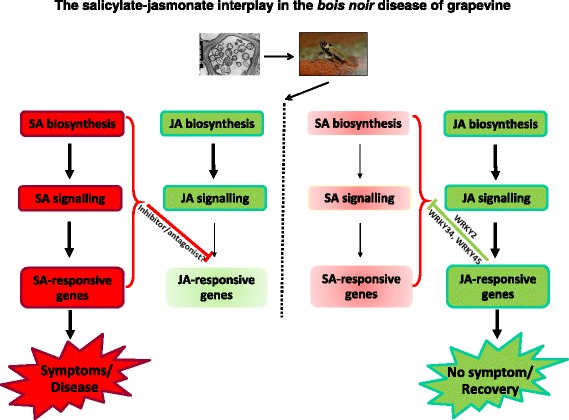
Results: Leaves from healthy, bois noir-diseased and bois noir-recovered plants were compared, both in the presence (late summer) and absence (late spring) of bois noir symptoms on the diseased plants. Analyses of salicylate and jasmonate contents, as well as the expression of genes involved in their biosynthesis, signalling and action, were evaluated. In symptomatic diseased plants (late summer), unlike symptomless plants (late spring), salicylate biosynthesis was increased and salicylate-responsive genes were activated. In contrast, jasmonate biosynthesis and signalling genes were up-regulated both in recovered and diseased plants at all sampling dates. The activation of salicylate signalling in symptomatic plants might have antagonised the jasmonate-mediated defence response by suppressing the expression of jasmonate-responsive genes.
Conclusions: Our results suggest that grapevine reacts to phytoplasma infection through salicylate-mediated signalling, although the resultant full activation of a salicylate-mediated response is apparently ineffective in conferring resistance against bois noir disease. Activation of the salicylate signalling pathway that is associated with the presence of bois noir phytoplasma seems to antagonise the jasmonate defence response, by failing to activate or suppressing both the expression of some jasmonate responsive genes that act downstream of the jasmonate biosynthetic pathway, as well as the first events of the jasmonate signalling pathway. On the other hand, activation of the entire jasmonate signalling pathway in recovered plants suggests the potential importance of jasmonate-regulated defences in preventing bois noir phytoplasma infections and the subsequent development of bois noir disease. Thus, on one hand, recovery could be achieved and maintained over time by preventing the activation of defence genes associated with salicylate signalling, and on the other hand, by activating jasmonate signalling and other defence responses.
Keywords: Bois noir disease and recovery; Grapevine; Jasmonate; Phytoplasmas; Plant-pathogen interactions; Salicylate; Stolbur; Vitis vinifera L.
Conflict of interest statement
Ethics approval and consent to participate
Samples were collected in a private vineyard, according to an informal agreement between the owner and the Plant Pathology scientists of the Department of Agricultural, Food, Environmental and Animal Sciences of the University of Udine, for monitoring this vineyard for
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
'Bois noir' phytoplasma induces significant reprogramming of the leaf transcriptome in the field grown grapevine.BMC Genomics. 2009 Oct 2;10:460. doi: 10.1186/1471-2164-10-460. BMC Genomics. 2009. PMID: 19799775 Free PMC article.
-
A successful defence strategy in grapevine cultivar 'Tocai friulano' provides compartmentation of grapevine Flavescence dorée phytoplasma.BMC Plant Biol. 2023 Mar 25;23(1):161. doi: 10.1186/s12870-023-04122-0. BMC Plant Biol. 2023. PMID: 36964496 Free PMC article.
-
Biochemical Changes in Leaves of Vitis vinifera cv. Sangiovese Infected by Bois Noir Phytoplasma.Pathogens. 2020 Apr 7;9(4):269. doi: 10.3390/pathogens9040269. Pathogens. 2020. PMID: 32272699 Free PMC article.
-
ORCAnization of jasmonate-responsive gene expression in alkaloid metabolism.Trends Plant Sci. 2001 May;6(5):212-9. doi: 10.1016/s1360-1385(01)01924-0. Trends Plant Sci. 2001. PMID: 11335174 Review.
-
Priming plant resistance by activation of redox-sensitive genes.Free Radic Biol Med. 2018 Jul;122:171-180. doi: 10.1016/j.freeradbiomed.2017.12.028. Epub 2017 Dec 24. Free Radic Biol Med. 2018. PMID: 29277443 Review.
Cited by
-
The Molecular Priming of Defense Responses is Differently Regulated in Grapevine Genotypes Following Elicitor Application against Powdery Mildew.Int J Mol Sci. 2020 Sep 15;21(18):6776. doi: 10.3390/ijms21186776. Int J Mol Sci. 2020. PMID: 32942781 Free PMC article.
-
Monoterpene Synthase Genes and Monoterpene Profiles in Pinus nigra subsp. laricio.Plants (Basel). 2022 Feb 6;11(3):449. doi: 10.3390/plants11030449. Plants (Basel). 2022. PMID: 35161430 Free PMC article.
-
Plant Hormones in Phytoplasma Infected Plants.Front Plant Sci. 2019 Apr 17;10:477. doi: 10.3389/fpls.2019.00477. eCollection 2019. Front Plant Sci. 2019. PMID: 31057582 Free PMC article. Review.
-
The Role of Plant Defense Signaling Pathways in Phytoplasma-Infected and Uninfected Aster Leafhoppers' Oviposition, Development, and Settling Behavior.J Chem Ecol. 2024 Jun;50(5-6):276-289. doi: 10.1007/s10886-024-01488-9. Epub 2024 Mar 26. J Chem Ecol. 2024. PMID: 38532167
-
'Candidatus Phytoplasma solani' interferes with the distribution and uptake of iron in tomato.BMC Genomics. 2019 Sep 10;20(1):703. doi: 10.1186/s12864-019-6062-x. BMC Genomics. 2019. PMID: 31500568 Free PMC article.
References
-
- Bertaccini A, Duduk B, Paltrinieri S, Contaldo N. Phytoplasmas and phytoplasma diseases: a severe threat to agriculture. Am J Plant Sci. 2014;5:1763–1788. doi: 10.4236/ajps.2014.512191. - DOI
-
- Belli G, Bianco PA, Conti M. Grapevine yellows in Italy: past, present and future. J Plant Pathol. 2010;92:303–326.
-
- Sforza R, Clair D, Daire X, Larrue J, Boudon-Padieu E. The role of Hyalesthes obsoletus (Hemiptera: Cixiidae) in the occurrence of bois noir of grapevines in France. J Phytopathol. 1998;146:549–556. doi: 10.1111/j.1439-0434.1998.tb04753.x. - DOI
-
- Osler R, Carraro L, Loi N, Refatti E. Symptom expression and disease occurrence of a yellows disease of grapevine in northeastern Italy. Plant Dis. 1993;77:496–498. doi: 10.1094/PD-77-0496. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
