

The Long Hunt for pssR-Looking for a Phospholipid Synthesis Transcriptional Regulator, Finding the Ribosome
- PMID: 28484043
- PMCID: PMC5494739
- DOI: 10.1128/JB.00202-17
The Long Hunt for pssR-Looking for a Phospholipid Synthesis Transcriptional Regulator, Finding the Ribosome
Abstract
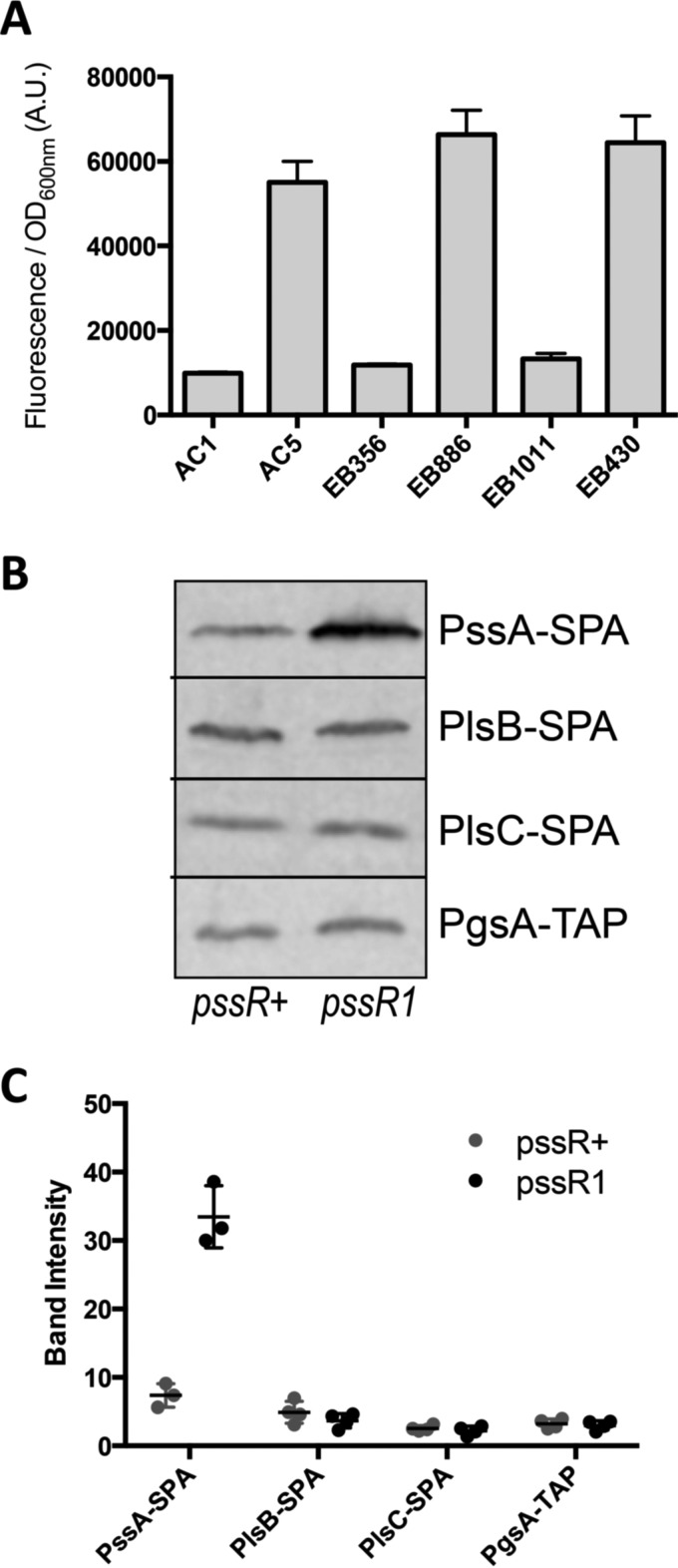
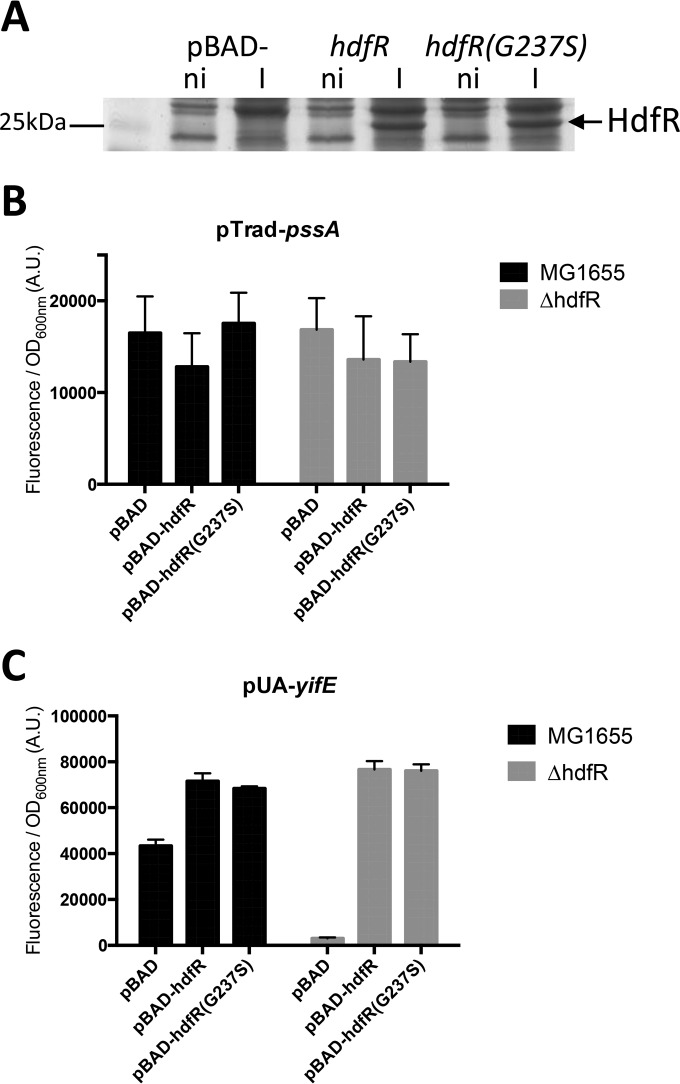
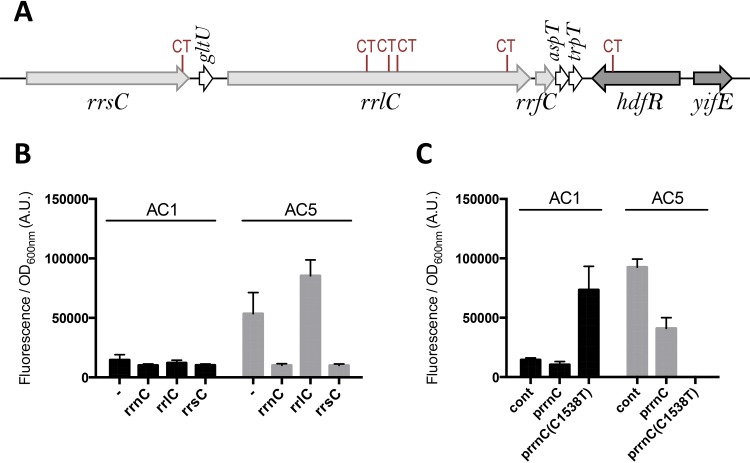
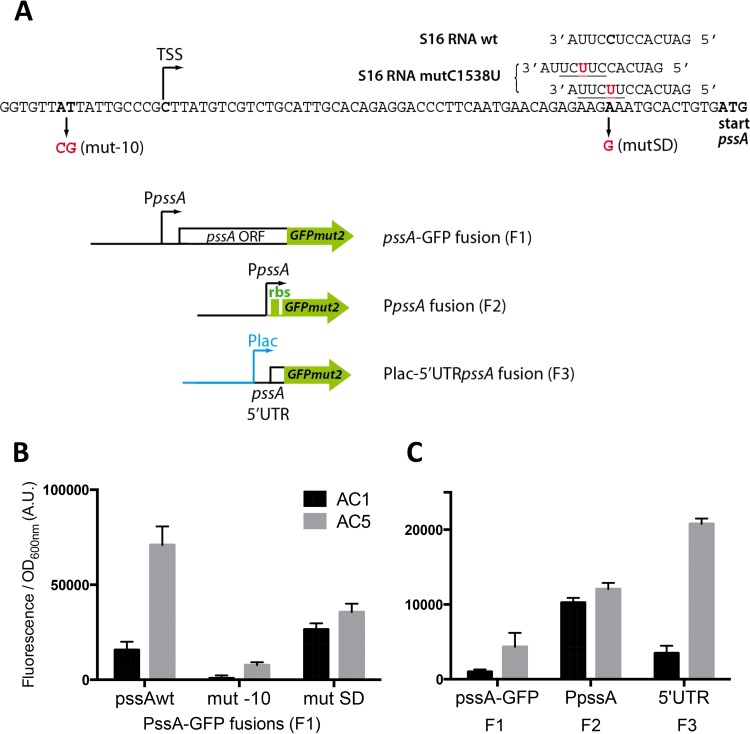
The phospholipid (PL) composition of bacterial membranes varies as a function of growth rate and in response to changes in the environment. While growth adaptation can be explained by biochemical feedback in the PL synthesis pathway, recent transcriptome studies have revealed that the expression of PL synthesis genes can also be tuned in response to various stresses. We previously showed that the BasRS two-component pathway controls the expression of the diacylglycerol kinase gene, dgkA, in Escherichia coli (A. Wahl, L. My, R. Dumoulin, J. N. Sturgis, and E. Bouveret, Mol Microbiol, 80:1260-1275, 2011, https://doi.org/10.1111/j.1365-2958.2011.07641.x). In this study, we set up a strategy to identify the mutation responsible for the upregulation of pssA observed in the historical pssR1 mutant and supposedly corresponding to a transcriptional repressor (C. P. Sparrow and J. Raetz, J Biol Chem, 258:9963-9967, 1983). pssA encodes phosphatidylserine synthase, the first step of phosphatidylethanolamine synthesis. We showed that this mutation corresponded to a single nucleotide change in the anti-Shine-Dalgarno sequence of the 16S rRNA encoded by the rrnC operon. We further demonstrated that this mutation enhanced the translation of pssA Though this effect appeared to be restricted to PssA among phospholipid synthesis enzymes, it was not specific, as evidenced by a global effect on the production of unrelated proteins.IMPORTANCE Bacteria adjust the phospholipid composition of their membranes to the changing environment. In addition to enzymatic regulation, stress response regulators control specific steps of the phospholipid synthesis pathway. We wanted to identify a potential regulator controlling the expression of the phosphatidylserine synthase gene. We showed that it was not the previously suggested hdfR gene and instead that a mutation in the anti-Shine-Dalgarno sequence of 16S RNA was responsible for an increase in pssA translation. This example underlines the fact that gene expression can be modulated by means other than specific regulatory processes.
Keywords: Escherichia coli; hdfR; maoP; phosphatidylserine synthase; phospholipid synthesis; pssA; pssR; ribosomal mutations; rrnC; yifE.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Antagonistic regulation of dgkA and plsB genes of phospholipid synthesis by multiple stress responses in Escherichia coli.Mol Microbiol. 2011 Jun;80(5):1260-75. doi: 10.1111/j.1365-2958.2011.07641.x. Epub 2011 Apr 5. Mol Microbiol. 2011. PMID: 21463370
-
A regulatory mechanism for the balanced synthesis of membrane phospholipid species in Escherichia coli.Biosci Biotechnol Biochem. 1996 Jan;60(1):111-6. doi: 10.1271/bbb.60.111. Biosci Biotechnol Biochem. 1996. PMID: 8824831
-
Dependency of sugar transport and phosphorylation by the phosphoenolpyruvate-dependent phosphotransferase system on membranous phosphatidylethanolamine in Escherichia coli: studies with a pssA mutant lacking phosphatidylserine synthase.Arch Microbiol. 2004 Jan;181(1):26-34. doi: 10.1007/s00203-003-0623-7. Epub 2003 Nov 21. Arch Microbiol. 2004. PMID: 14634719
-
Specialized ribosomes in Escherichia coli.Biotechnol Prog. 1993 Sep-Oct;9(5):443-9. doi: 10.1021/bp00023a001. Biotechnol Prog. 1993. PMID: 7764160 Review.
-
Regulation of arginine biosynthesis, catabolism and transport in Escherichia coli.Amino Acids. 2019 Aug;51(8):1103-1127. doi: 10.1007/s00726-019-02757-8. Epub 2019 Jul 3. Amino Acids. 2019. PMID: 31267155 Review.
Cited by
-
Pseudomonas aeruginosa adapts to octenidine via a combination of efflux and membrane remodelling.Commun Biol. 2021 Sep 9;4(1):1058. doi: 10.1038/s42003-021-02566-4. Commun Biol. 2021. PMID: 34504285 Free PMC article.
-
Identifying Small Proteins by Ribosome Profiling with Stalled Initiation Complexes.mBio. 2019 Mar 5;10(2):e02819-18. doi: 10.1128/mBio.02819-18. mBio. 2019. PMID: 30837344 Free PMC article.
-
A systematic analysis of hypermucoviscosity and capsule reveals distinct and overlapping genes that impact Klebsiella pneumoniae fitness.PLoS Pathog. 2021 Mar 15;17(3):e1009376. doi: 10.1371/journal.ppat.1009376. eCollection 2021 Mar. PLoS Pathog. 2021. PMID: 33720976 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
