

Insights into the Structure, Function, and Ion-Mediated Signaling Pathways Transduced by Plant Integrin-Linked Kinases
- PMID: 28421082
- PMCID: PMC5376563
- DOI: 10.3389/fpls.2017.00376
Insights into the Structure, Function, and Ion-Mediated Signaling Pathways Transduced by Plant Integrin-Linked Kinases
Abstract
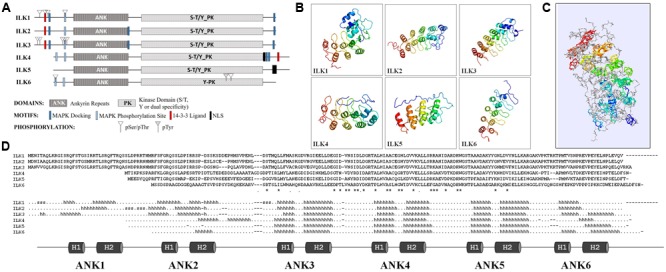
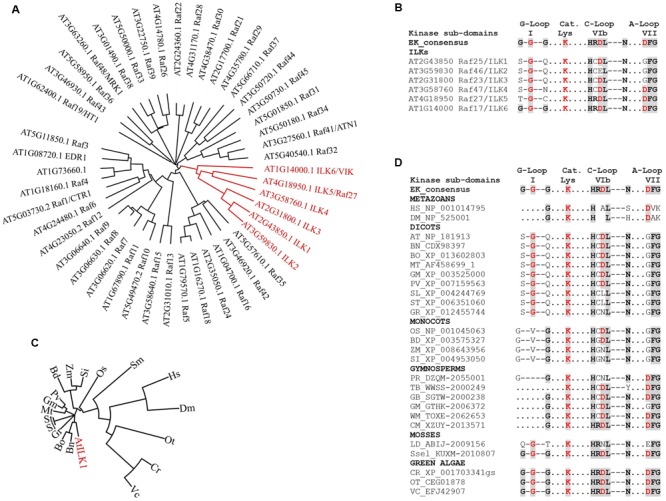
Kinases facilitate detection of extracellular signals and set in motion cellular responses for plant adaptation and survival. Some of the energy utilized for kinase signal processing is produced through the activity of ion transporters. Additionally, the synergy between cellular ions and signal transduction influences plant response to pathogens, and their growth and development. In plants, the signaling elements that connect cell wall and membrane sensors with ion homeostasis and transport-mediated processes are largely unknown. Current research indicates that plant Integrin-Linked Kinases (ILKs), a subfamily Raf-like MAP2K Kinases, may have evolved to fulfill this role. In this review, we explore new findings on plant ILKs placing a particular focus on the connection between ILKs proteins unique structural features and ILKs functions. The ankyrin repeat motifs and the kinase domains of ILKs in Arabidopsis and land plants lineage, respectively, are analyzed and discussed as potential determinants of ILKs' metal ion cofactor specificity and their enzymatic and interaction activities. Further, ILKs regulation through gene expression, subcellular localization, and ions and ion transporters is reviewed in the context of recent studies. Finally, using evidence from literature and interactomics databanks, we infer ILKs-dependent cellular pathways and highlight their potential in transmitting multiple types of signals originating at the interface between the cell wall and plasma membrane.
Keywords: calcium; integrin-linked kinases; manganese; plant abiotic response; plant immune response; potassium; transporters.
Figures

Similar articles
-
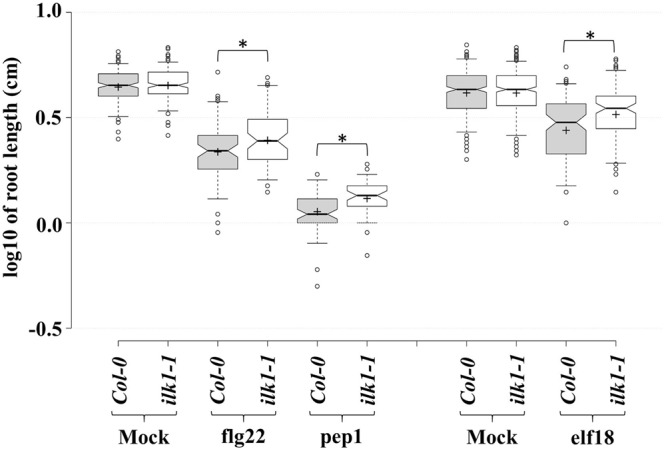
The Raf-like Kinase ILK1 and the High Affinity K+ Transporter HAK5 Are Required for Innate Immunity and Abiotic Stress Response.Plant Physiol. 2016 Jun;171(2):1470-84. doi: 10.1104/pp.16.00035. Epub 2016 May 2. Plant Physiol. 2016. PMID: 27208244 Free PMC article.
-
A mutant ankyrin protein kinase from Medicago sativa affects Arabidopsis adventitious roots.Funct Plant Biol. 2008 Feb;35(1):92-101. doi: 10.1071/FP07209. Funct Plant Biol. 2008. PMID: 32688760
-
Insights on Calcium-Dependent Protein Kinases (CPKs) Signaling for Abiotic Stress Tolerance in Plants.Int J Mol Sci. 2019 Oct 24;20(21):5298. doi: 10.3390/ijms20215298. Int J Mol Sci. 2019. PMID: 31653073 Free PMC article. Review.
-
Molecular insights into the role of plant transporters in salt stress response.Physiol Plant. 2021 Dec;173(4):1481-1494. doi: 10.1111/ppl.13453. Epub 2021 May 28. Physiol Plant. 2021. PMID: 33963568 Review.
-
Structural basis of the regulatory mechanism of the plant CIPK family of protein kinases controlling ion homeostasis and abiotic stress.Proc Natl Acad Sci U S A. 2014 Oct 21;111(42):E4532-41. doi: 10.1073/pnas.1407610111. Epub 2014 Oct 6. Proc Natl Acad Sci U S A. 2014. PMID: 25288725 Free PMC article.
Cited by
-
Insights into the effect of human civilization on Malus evolution and domestication.Plant Biotechnol J. 2021 Nov;19(11):2206-2220. doi: 10.1111/pbi.13648. Epub 2021 Jul 7. Plant Biotechnol J. 2021. PMID: 34161653 Free PMC article.
-
Primary nitrate responses mediated by calcium signalling and diverse protein phosphorylation.J Exp Bot. 2020 Jul 25;71(15):4428-4441. doi: 10.1093/jxb/eraa047. J Exp Bot. 2020. PMID: 31985788 Free PMC article. Review.
-
The Ankyrin-Repeat Gene GmANK114 Confers Drought and Salt Tolerance in Arabidopsis and Soybean.Front Plant Sci. 2020 Oct 29;11:584167. doi: 10.3389/fpls.2020.584167. eCollection 2020. Front Plant Sci. 2020. PMID: 33193533 Free PMC article.
-
Comprehensive transcriptome analyses of Fusarium-infected root xylem tissues to decipher genes involved in chickpea wilt resistance.3 Biotech. 2023 Dec;13(12):390. doi: 10.1007/s13205-023-03803-9. Epub 2023 Nov 7. 3 Biotech. 2023. PMID: 37942053 Free PMC article.
-
Towards engineering ectomycorrhization into switchgrass bioenergy crops via a lectin receptor-like kinase.Plant Biotechnol J. 2021 Dec;19(12):2454-2468. doi: 10.1111/pbi.13671. Epub 2021 Aug 11. Plant Biotechnol J. 2021. PMID: 34272801 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous